Synthetic pesticides present risks of pollution of the environment, humans and livestock and the alternative proposed today is to use botanical extracts in the fields against crop pests. But in North Cameroon, little information exists concerning the effect of these extracts on useful pollinating insects in general and no information exists in particular on foragers of the genus Amegilla Friese, 1897 (Apidae: Apinae: Anthophorini). The frequency and foraging activities of Amegilla, on newly blooming flowers of Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae) were recorded during five consecutive days in 2021 and 2022 planting campaigns. Plants were divided into untreated plots and plots treated using the synthetic insecticide Parastar (l p.c..ha-1) or 10%, 20% and 30% aqueous leaf extracts of Calotropis procera (Aiton) Aiton, 1811 (Gentianales: Apocynaceae), Eucalyptus camaldulensis Dehnh., 1832 (Myrtales: Myrtaceae) and Tithonia diversifolia (Hemsley) Gray, 1883 (Asterales: Asteraceae) respectively. Among 8,987 insects collected (48.9% in 2021), Amegila calens Le Peletier. 1841 with stockier foragers (2021 campaign: 2.2% of the total collection, entomophily FA. calens=4.5%; 2022 campaign: 0.7%, FA. calens=1.3%; pooled campaigns: 2.9%, FA. calens=2.9%) and Amegilla sp. with slender foragers (2021: 3.8%, FAmegilla sp.=7.7%; 2022: no data) were recorded. Foragers started activity from 6 a.m. and stopped foraging before noon, with a peak of activity in 8 to 9 a.m. time slot for A. calens and 10 to 11 a.m. time slot for Amegilla sp.. During the five consecutive days from the first blooming day of the flowers, 598 visits (89.8% in 2021 and 10.2% in 2022) were recorded with a peak of visits during the 3rd day and then declined until it stopped during the 5th day. Treatments including the synthetic insecticide (which was the most repellent to the wild bees), did not significantly reduce the frequency of visits. But 20% aqueous extract of Ca. procera showed a significant increased of the mean duration of visits of the bees, compare to the results recorded in Parastar-treated plots. Therefore, the tested extracts, especially 20% aqueous leaves extract of Ca. procera may be recommended to control field insect pests and for preservation of foraging activities of Amegilla genus.
| Published in | American Journal of Entomology (Volume 8, Issue 3) |
| DOI | 10.11648/j.aje.20240803.13 |
| Page(s) | 76-101 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2024. Published by Science Publishing Group |
Wild Bees, Vigna unguiculata, Synthetic Insecticide, Leaves Extract, Inhibition Effect, Dang
A1 | F1 | A1,000=(A1/F1)*1000 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
n | Min. | Max. | Mean±se | Min. | Max. | Mean±se | Min. | Max. | Mean±se | |
A. Pooled years and Amegilla species | ||||||||||
Untreated | 145 | 1 | 8 | 3±0 | 5 | 689 | 222±16 | 2 | 800 | 24±6 |
Parastar | 61 | 1 | 7 | 2±0 | 5 | 640 | 208±22 | 2 | 400 | 25±7 |
Cp10 | 109 | 1 | 8 | 3±0 | 5 | 689 | 250±18 | 1 | 800 | 24±7 |
Cp20 | 101 | 1 | 8 | 3±0 | 15 | 689 | 256±20 | 1 | 179 | 19±2 |
Cp30 | 56 | 1 | 8 | 3±0 | 15 | 689 | 249±28 | 1 | 133 | 21±3 |
Ec10 | 125 | 1 | 9 | 3±0 | 5 | 650 | 212±16 | 2 | 800 | 28±7 |
Ec20 | 117 | 1 | 9 | 2±0 | 5 | 689 | 218±18 | 1 | 800 | 27±7 |
Ec30 | 54 | 1 | 7 | 3±0 | 25 | 689 | 251±29 | 1 | 100 | 20±3 |
Td10 | 102 | 1 | 8 | 3±0 | 24 | 689 | 261±20 | 1 | 179 | 17±2 |
Td20 | 91 | 1 | 8 | 3±0 | 24 | 689 | 286±23 | 1 | 87 | 17±2 |
Td30 | 46 | 1 | 8 | 3±0 | 24 | 650 | 222±30 | 3 | 42 | 17±2 |
Pooled plots | 1007 | 1 | 9 | 3±0 | 5 | 689 | 239±6 | 1 | 800 | 22±2 |
ANOVA | F(10; 996)=1.013, p=0.430 ns | F(10; 996)=1.463, p=0.148 ns | F(10; 996)=5.238, p=0.874 ns | |||||||
B. Pooled Amegilla in 2021 | ||||||||||
Untreated | 95 | 1 | 8 | 2±0 | 15 | 689 | 198±18 | 2 | 80 | 17±1 |
Parastar | 37 | 1 | 7 | 2±0 | 5 | 630 | 237±27 | 2 | 400 | 23±11 |
Cp10 | 71 | 1 | 8 | 3±0 | 24 | 650 | 238±23 | 4 | 179 | 19±3 |
Cp20 | 65 | 1 | 7 | 3±0 | 15 | 650 | 219±24 | 3 | 179 | 21±3 |
Cp30 | 36 | 1 | 8 | 3±0 | 100 | 689 | 342±31 | 1 | 59 | 13±2 |
Ec10 | 83 | 1 | 9 | 2±0 | 15 | 650 | 211±21 | 3 | 179 | 20±3 |
Ec20 | 77 | 1 | 9 | 2±0 | 15 | 650 | 194±21 | 3 | 179 | 22±3 |
Ec30 | 35 | 1 | 7 | 3±0 | 100 | 689 | 330±34 | 1 | 59 | 13±2 |
A1 | F1 | A1,000=(A1/F1)*1000 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
n | Min. | Max. | Mean±se | Min. | Max. | Mean±se | Min. | Max. | Mean±se | |
B. Pooled Amegilla in 2021 (Continued) | ||||||||||
Td10 | 65 | 1 | 8 | 3±0 | 24 | 650 | 241±26 | 6 | 179 | 20±3 |
Td20 | 60 | 1 | 8 | 3±0 | 24 | 650 | 255±27 | 3 | 87 | 17±2 |
Td 30 | 30 | 1 | 8 | 3±0 | 35 | 650 | 276±42 | 3 | 40 | 14±2 |
Pooled plots | 654 | 1 | 9 | 3±0 | 5 | 689 | 236±8 | 1 | 400 | 19±1 |
ANOVA | F(10; 643)=2.726, p=0.003 * | F(10; 643)=3.008, p=0.001 * | F(10; 643)=0.864, p=0.567 ns | |||||||
C. Amegilla calens in 2021 | ||||||||||
Untreated | 60 | 1 | 6 | 2±0 | 15 | 600 | 119±15 | 3 | 80 | 21±2 |
Parastar | 20 | 1 | 6 | 2±0 | 5 | 630 | 185±29 | 2 | 400 | 32±19 |
Cp 10 | 43 | 1 | 6 | 2±0 | 48 | 650 | 174±23 | 6 | 69 | 16±2 |
Cp 20 | 40 | 1 | 6 | 2±0 | 15 | 650 | 168±24 | 3 | 69 | 18±3 |
Cp 30 | 22 | 1 | 8 | 4±0 | 100 | 689 | 351±40 | 2 | 30 | 11±1 |
Ec 10 | 53 | 1 | 6 | 2±0 | 15 | 650 | 160±22 | 3 | 69 | 19±2 |
Ec 20 | 50 | 1 | 6 | 2±0 | 15 | 650 | 155±23 | 3 | 69 | 19±2 |
Ec 30 | 22 | 1 | 7 | 3±0 | 100 | 650 | 335±40 | 4 | 30 | 11±1 |
Td 10 | 39 | 1 | 6 | 2±0 | 48 | 650 | 160±24 | 6 | 69 | 17±2 |
Td 20 | 35 | 1 | 6 | 2±0 | 48 | 650 | 171±27 | 3 | 69 | 16±2 |
Td 30 | 18 | 1 | 6 | 2±0 | 48 | 650 | 172±41 | 3 | 40 | 14±2 |
Pooled plots | 402 | 1 | 8 | 2±0 | 5 | 689 | 178±8 | 2 | 400 | 18±1 |
ANOVA | F(10; 391)=6.464, p<0.001 * | F(10; 391)=6.147, p<0.001 * | F(10; 391)=1.249, p=0.258 ns | |||||||
D. Amegilla sp. in 2021 | ||||||||||
Untreated | 35 | 1 | 8 | 3±0 | 100 | 689 | 333±32 | 2 | 22 | 10±1 |
Parastar | 17 | 1 | 7 | 3±0 | 100 | 630 | 299±45 | 2 | 35 | 13±3 |
Cp 10 | 28 | 1 | 8 | 4±0 | 24 | 650 | 337±42 | 4 | 179 | 24±7 |
Cp 20 | 25 | 1 | 7 | 4±0 | 24 | 650 | 302±43 | 4 | 179 | 25±7 |
Cp 30 | 14 | 1 | 8 | 3±1 | 102 | 689 | 326±53 | 1 | 59 | 16±6 |
Ec 10 | 30 | 1 | 9 | 3±0 | 28 | 650 | 303±38 | 4 | 179 | 22±6 |
Ec 20 | 27 | 1 | 9 | 3±0 | 24 | 640 | 267±39 | 4 | 179 | 28±7 |
Ec 30 | 13 | 1 | 7 | 3±1 | 102 | 689 | 322±65 | 1 | 59 | 16±5 |
Td 10 | 26 | 1 | 8 | 4±0 | 24 | 650 | 362±45 | 6 | 179 | 25±7 |
Td 20 | 25 | 1 | 8 | 5±0 | 24 | 650 | 372±41 | 6 | 87 | 19±3 |
Td 30 | 12 | 1 | 8 | 5±1 | 35 | 650 | 431±63 | 6 | 29 | 14±2 |
Pooled plots | 252 | 1 | 9 | 4±0 | 24 | 689 | 328±13 | 1 | 179 | 20±2 |
ANOVA | F(10; 241)=2.680, p=0.004 * | F(10; 241)=0.833, p=0.597 ns | F(10; 241)=1.093, p=0.368 ns | |||||||
E. Amegilla calens in 2022 | ||||||||||
Untreated | 50 | 1 | 8 | 4±0 | 5 | 650 | 267±28 | 2 | 800 | 37±16 |
Parastar | 24 | 1 | 6 | 2±0 | 25 | 640 | 164±37 | 2 | 100 | 28±5 |
A1 | F1 | A1,000=(A1/F1)*1000 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
n | Min. | Max. | Mean±se | Min. | Max. | Mean±se | Min. | Max. | Mean±se | |
E. Amegilla calens in 2022 (Continued) | ||||||||||
Cp 10 | 38 | 1 | 7 | 2±0 | 5 | 689 | 273±28 | 1 | 800 | 33±21 |
Cp 20 | 36 | 1 | 8 | 3±0 | 29 | 689 | 323±36 | 1 | 79 | 15±3 |
Cp 30 | 20 | 1 | 6 | 2±0 | 15 | 600 | 83±28 | 2 | 133 | 35±7 |
Ec 10 | 42 | 1 | 9 | 3±0 | 5 | 630 | 214±26 | 2 | 800 | 44±19 |
Ec 20 | 40 | 1 | 8 | 3±0 | 5 | 689 | 264±31 | 1 | 800 | 36±20 |
Ec 30 | 19 | 1 | 6 | 2±0 | 25 | 600 | 105±33 | 2 | 100 | 32±6 |
Td 10 | 37 | 1 | 7 | 2±0 | 100 | 689 | 296±31 | 1 | 36 | 11±2 |
Td 20 | 31 | 1 | 8 | 3±0 | 29 | 689 | 347±41 | 1 | 79 | 17±4 |
Td 30 | 16 | 1 | 8 | 2±1 | 24 | 300 | 121±22 | 7 | 42 | 22±3 |
Pooled plots | 353 | 1 | 9 | 3±0 | 5 | 689 | 244±10 | 1 | 800 | 29±5 |
ANOVA | F(10; 342)=2.497, p=0.007 * | F(10; 342)=6.020, p<0.001 * | F(10; 342)=0.525, p=0.872 ns | |||||||
A1: pooled Amegilla in 2021: | Untreated vs. Cp30: | p=0.023 *; | Other comparisons were not significant | |||
A1: A. calens in 2021: | Untreated vs. Cp30: | p=1x10-5 *; | Cp20 vs. Cp30: | p=4x10-5 *; | Cp30 vs. Td30: | p=1.6x10-4 * |
Untreated vs. Ec30: | p=1x10-5 * | Cp20 vs. Ec30: | p=5.5x10-4 *; | Ec 30 vs. Ec 10: | p=4.2x10-4 * | |
Parastar vs. Ec30: | p=0.006 * | Cp30 vs. Ec10: | p=4x10-5 *; | Ec 30 vs. Ec 20: | p=3.6x10-4 * | |
Parastar vs. Cp30: | p=6.5x10-4 *; | Cp30 vs. Ec20: | p=2x10-5 * | Ec 30 vs. Td 10: | p=6.8x10-4 * | |
Cp10 vs. Cp30: | p=2.1x10-4 *; | Cp30 vs. Td10: | p=6x10-5 *; | Ec 30 vs. Td 20: | p=0.001 * | |
Cp10 vs. Ec30: | p=0.001 * | Cp30 vs. Td20: | p=8x10-5 *; | Ec 30 vs. Td 30: | p=0.002 * | |
Other comparisons were not significant | ||||||
A1: Amegilla sp. in 2021: | Parastar vs. Td20: | p=0.029 * | Other comparisons were not significant | |||
A1: A. calens in 2022: | Untreated vs. Cp10: | p=0.045 *; | Untreated vs. Cp30: | p=0.019 *; | ||
Other comparisons were not significant | ||||||
F1: pooled Amegilla in 2021: | Untreated vs. Cp30: | p=0.006 *; | Cp30 vs. Ec10: | p=0.021 * | Ec10 vs. Ec30: | p=0.047 * |
Untreated vs. Ec30: | p=0.016 * | Cp30 vs. Ec20: | p= 0.007 *; | Ec20 vs. Ec30: | p=0.020 *; | |
Cp20 vs. Cp30: | p=0.048 *; | Other comparisons were not significant | ||||
F1: A. calens in 2021: | Untreated vs. Cp30: | p=1x10-5 *; | Cp20 vs. Cp30: | p=1.8x10-4 *; | Cp30 vs. Td30: | p=0.003 * |
Untreated vs. Ec30: | p=1x10-5 * | Cp20 vs. Ec30: | p=6.8x10-4 *; | Ec10 vs. Ec30: | p=2.4x10-4 * | |
Parastar vs. Cp30: | p=0.001 *; | Cp30 vs. Ec10: | p=5x10-5 *; | Ec20 vs. Ec30: | p=2.1x10-4 * | |
Parastar vs. Ec30: | p=0.002 * | Cp30 vs. Ec20: | p=4x10-5 * | Ec 30 vs. Td 10: | p=4.7x10-4 * | |
Cp10 vs. Cp30: | p=8x10-5 *; | Cp30 vs. Td10: | p=1.3x10-4 *; | Ec 30 vs. Td 20: | p=9.3x10-4 * | |
Cp10 vs. Ec30: | p=2x10-4 * | Cp30 vs. Td20: | p=2.8x10-4 *; | Ec 30 vs. Td 30: | p=0.005 * | |
Other comparisons were not significant | ||||||
F1: A. calens in 2022: | Untreated vs. Cp30: | p=0.015 *; | Cp20 vs. Cp30: | p=1.4x10-4 *; | Ec10 vs. Td20: | p=0.039 *; |
Untreated vs. Ec30: | p=0.015 *; | Cp20 vs. Ec30: | p=0.001 *; | Ec20 vs. Ec30: | p=0.016 * | |
Untreated vs. Td30: | p=0.046 *; | Cp20 vs. Td30: | p=0.006 *; | Ec20 vs. Td30: | p=0.042 * | |
Parastar vs. Cp20: | p=0.019 *; | Cp30 vs. Ec20: | p=0.004 * | Ec 30 vs. Td10: | p=0.006 * | |
Parastar vs. Td20: | p=0.007 *; | Cp30 vs. Td10: | p=0.001 *; | Ec30 vs. Td20: | p=3.0x10-4 * | |
Cp10 vs. Cp30: | p=0.005 *; | Cp30 vs. Td20: | p=4.0x10-5 *; | Td20 vs. Td30: | p=0.002 * | |
Cp10 vs. Ec30: | p=0.020 *; | Td10 vs. Td30: | p=0.025 *; | Other comparisons not significant | ||
A. Pooled species in the pooled years | B. Amegilla calens in the pooled years | C. A. calens in 2021 | ||||||||||||
Plots | n | Min. | Max. | Mean±se | n | Min. | Max. | Mean±se | n | Min. | Max. | Mean±se | ||
Unt. | 158 | 1.0 | 9.0 | 4.0±0.2 | 109 | 1.0 | 9.0 | 4.5±0.2 | 44 | 1.0 | 8.0 | 3.1±0.3 | ||
Para. | 77 | 1.0 | 9.0 | 3.3±0.3 | 67 | 1.0 | 9.0 | 3.6±0.3 | 24 | 1.0 | 3.0 | 1.5±0.1 | ||
Cp10 | 132 | 1.0 | 9.0 | 4.1±0.2 | 96 | 1.0 | 9.0 | 4.4±0.2 | 44 | 2.0 | 8.0 | 3.6±0.2 | ||
Cp20 | 137 | 1.0 | 9.0 | 4.5±0.2 | 96 | 1.0 | 9.0 | 5.1±0.2 | 41 | 1.0 | 6.0 | 3.8±0.2 | ||
Cp30 | 90 | 1.0 | 9.0 | 3.8±0.2 | 67 | 1.0 | 9.0 | 4.2±0.3 | 35 | 2.0 | 7.0 | 3.5±0.3 | ||
Ec10 | 142 | 1.0 | 9.0 | 4.1±0.2 | 100 | 1.0 | 9.0 | 4.5±0.2 | 43 | 1.0 | 6.0 | 3.3±0.3 | ||
Ec20 | 144 | 1.0 | 9.0 | 3.7±0.2 | 98 | 1.0 | 9.0 | 4.4±0.3 | 40 | 1.0 | 7.0 | 2.5±0.3 | ||
Ec30 | 102 | 1.0 | 8.0 | 3.4±0.2 | 74 | 1.0 | 8.0 | 3.9±0.2 | 34 | 1.0 | 7.0 | 3.0±0.3 | ||
Td10 | 105 | 1.0 | 9.0 | 3.4±0.2 | 75 | 1.0 | 9.0 | 4.0±0.3 | 30 | 1.0 | 7.0 | 2.5±0.3 | ||
Td20 | 133 | 1.0 | 9.0 | 3.6±0.2 | 93 | 1.0 | 9.0 | 4.1±0.2 | 41 | 1.0 | 5.0 | 2.5±0.2 | ||
Td30 | 72 | 1.0 | 8.0 | 3.7±0.3 | 56 | 1.0 | 8.0 | 4.3±0.3 | 27 | 2.0 | 8.0 | 3.5±0.3 | ||
Pooled plots | 1,292 | 1.0 | 9.0 | 3.8±0.1 | 931 | 1.0 | 9.0 | 4.30±0.07 | 403 | 1.0 | 8.0 | 3.06±0.08 | ||
ANOVA | F(10; 1281)=3.043, p=8x10-5 * | F(10; 920)=2.534, p=0.005 * | F(10; 392)=5.708, p<0.001 * | |||||||||||
Unt. vs. Para | Student test: t=2.326, df=233, p=0.021* | Student test: t=2.731, df=174, p=0.007 * | Student test: t=4.205, df=66, p=8.0x10-5 * | |||||||||||
D. Amegilla sp. in 2021 | E. A. calens in 2022 | |||||||||||||
Plots | n | Min. | Max. | Mean±se | n | Min. | Max. | Mean±se | ||||||
Unt. | 49 | 1.0 | 8.0 | 2.8±0.2 | 65 | 1.0 | 9.0 | 5.5±0.3 | ||||||
Para. | 10 | 1.0 | 2.0 | 1.3±0.2 | 43 | 2.0 | 9.0 | 4.7±0.3 | ||||||
Cp10 | 36 | 2.0 | 8.0 | 3.4±0.3 | 52 | 1.0 | 9.0 | 5.0±0.3 | ||||||
Cp20 | 41 | 1.0 | 6.0 | 3.1±0.2 | 55 | 2.0 | 9.0 | 6.0±0.2 | ||||||
Cp30 | 23 | 2.0 | 7.0 | 2.7±0.3 | 32 | 1.0 | 9.0 | 5.0±0.4 | ||||||
Ec10 | 42 | 1.0 | 6.0 | 3.2±0.3 | 57 | 2.0 | 9.0 | 5.3±0.2 | ||||||
D. Amegilla sp. in 2021 | E. A. calens in 2022 | |||||||
|---|---|---|---|---|---|---|---|---|
Plots | n | Min. | Max. | Mean±se | n | Min. | Max. | Mean±se |
Ec20 | 46 | 1.0 | 7.0 | 2.4±0.2 | 58 | 1.0 | 9.0 | 5.6±0.3 |
Ec30 | 28 | 1.0 | 7.0 | 2.3±0.3 | 40 | 1.0 | 8.0 | 4.6±0.4 |
Td10 | 30 | 1.0 | 5.0 | 2.1±0.2 | 45 | 1.0 | 9.0 | 4.9±0.3 |
Td20 | 40 | 1.0 | 5.0 | 2.4±0.2 | 52 | 1.0 | 9.0 | 5.3±0.3 |
Td30 | 16 | 1.0 | 4.0 | 1.8±0.2 | 29 | 1.0 | 8.0 | 5.0±0.4 |
Pooled plots | 361 | 1.0 | 8.0 | 2.66±0.08 | 528 | 1.0 | 9.0 | 5.2±0.1 |
ANOVA | F(10; 350)=3.956, p=4x10-5 * | F(10; 517)=1.786, p=0.060 ns | ||||||
Unt. vs. Para. | Student test: t=2.856, df=57, p=0.006 * | Student test: t=1.882, df=106, p=0.063 ns | ||||||
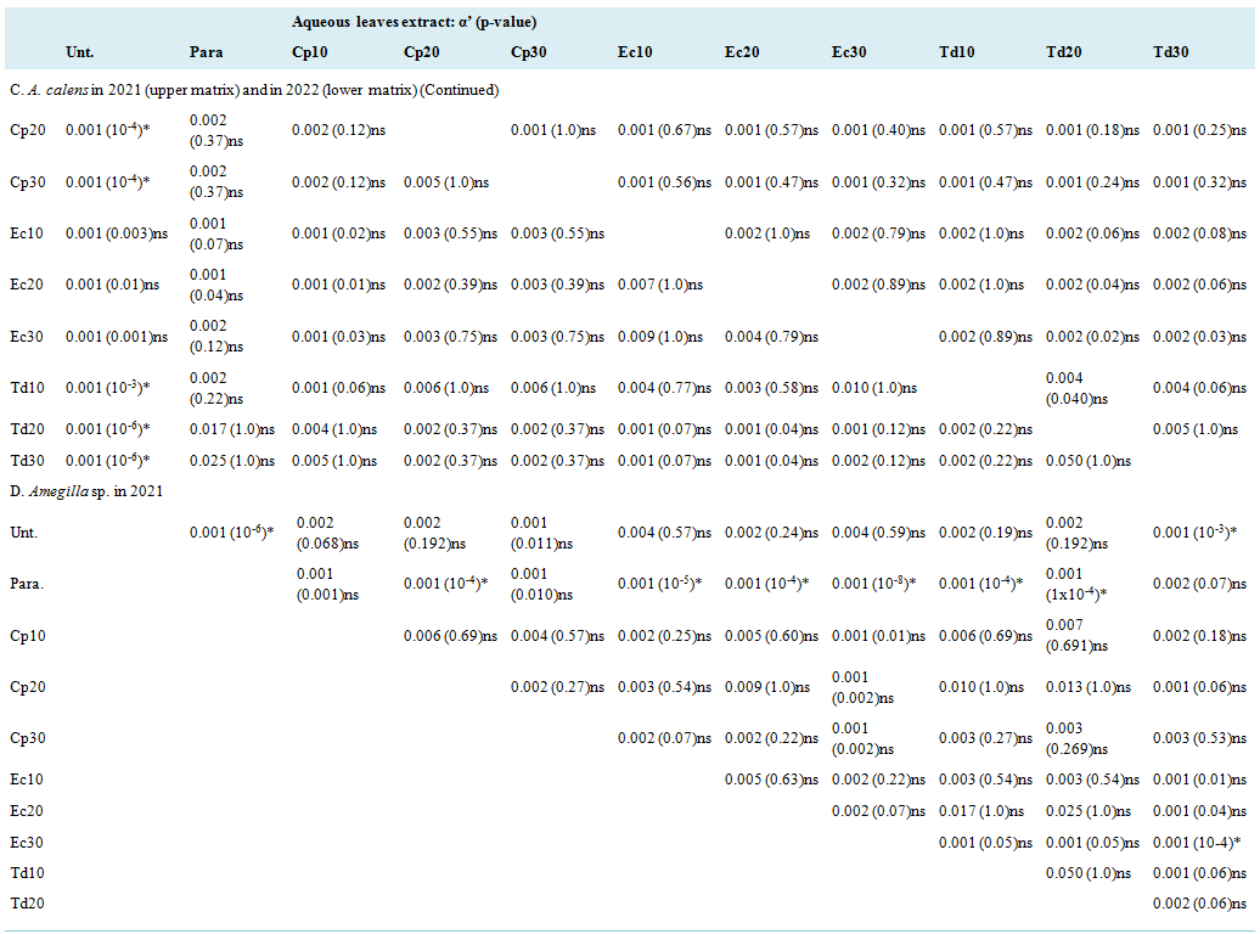
Aqueous leaves extract: p-value | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Unt. | Para | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
A. Pooled species in the pooled years (upper part matrix); Amegilla calens in the pooled years (lower part matrix) | |||||||||||
Unt. | 0.899ns | 0.249 ns | 0.591ns | 0.739ns | 0.549ns | 0.339ns | 0.392ns | 0.515ns | 0.786ns | ||
Parastar | 0.184ns | 0.006 * | 0.630ns | 0.180ns | 0.667ns | 0.867ns | 0.615ns | 0.736ns | 0.745ns | ||
Cp10 | 0.901ns | 0.281ns | 0.177 ns | 0.806ns | 0.913ns | 0.609ns | 0.279ns | 0.309ns | 0.461ns | 0.806ns | |
Cp20 | 0.091ns | 0.001 * | 0.929ns | 0.213ns | 0.298ns | 0.054ns | 0.010 * | 0.010 * | 0.022 * | 0.198ns | |
Cp30 | 0.959ns | 0.413ns | 0.972ns | 0.240ns | 0.695ns | 0.713ns | 0.709ns | 0.791ns | 0.837ns | 0.919ns | |
Ec10 | 0.791ns | 0.234ns | 0.866ns | 0.135ns | 0.976ns | 0.535ns | 0.264ns | 0.299ns | 0.431ns | 0.760ns | |
Ec20 | 0.952ns | 0.268ns | 0.166ns | 0.195ns | 0.930ns | 0.964ns | 0.726ns | 0.828ns | 0.852ns | 0.930ns | |
Ec30 | 0.588ns | 0.410ns | 0.750ns | 0.022 * | 0.776ns | 0.711ns | 0.717ns | 0.992ns | 0.597ns | 0.708ns | |
Td10 | 0.686ns | 0.550ns | 0.806ns | 0.038 * | 0.740ns | 0.785ns | 0.760ns | 0.824ns | 0.849ns | 0.852ns | |
Td20 | 0.815ns | 0.453ns | 0.890ns | 0.061ns | 0.693ns | 0.884ns | 0.838ns | 0.806ns | 0.694ns | 0.722ns | |
Td30 | 0.940ns | 0.543ns | 0.921ns | 0.259ns | 0.978ns | 0.951ns | 0.755ns | 0.883ns | 0.884ns | 0.914ns | |
B. A. calens in 2021 (upper part matrix); Amegilla sp. in 2021 (lower part matrix) | |||||||||||
Unt. | 0.566ns | 0.408ns | 0.712ns | 0.574ns | 0.384ns | 0.765ns | 0.366ns | 0.183ns | 0.641ns | ||
Parastar | 2x10-5* | 2x10-5 * | 5x10-5* | 1x10-4* | 0.011ns | 0.002* | 0.043 * | 0.050ns | 2x10-4 * | ||
Cp10 | 0.340ns | 0.005 * | 0.672ns | 0.731ns | 0.792ns | 0.033ns | 0.536ns | 0.060ns | 0.021 * | 0.914ns | |
Cp20 | 0.371ns | 0.018 * | 0.718ns | 0.742ns | 0.674ns | 0.012 * | 0.375ns | 0.026 * | 0.008 * | 0.869ns | |
Cp30 | 0.748ns | 0.167ns | 0.448ns | 0.553ns | 0.858ns | 0.116ns | 0.702ns | 0.155ns | 0.074ns | 0.935ns | |
Ec10 | 0.411ns | 0.010 * | 0.664ns | 0.721ns | 0.534ns | 0.185ns | 0.689ns | 0.211ns | 0.097ns | 0.686ns | |
Ec20 | 0.504ns | 0.218ns | 0.053ns | 0.175ns | 0.702ns | 0.099ns | 0.513ns | 0.982ns | 0.999ns | 0.179ns | |
Ec30 | 0.558ns | 0.273ns | 0.079ns | 0.225ns | 0.761ns | 0.138ns | 0.767ns | 0.417ns | 0.176ns | 0.679ns | |
Td10 | 0.353ns | 0.275ns | 0.024 * | 0.098ns | 0.650ns | 0.048 * | 0.740ns | 0.697ns | 0.993ns | 0.203ns | |
Td20 | 0.432ns | 0.267ns | 0.067ns | 0.175ns | 0.487ns | 0.114ns | 0.917ns | 0.923ns | 0.849ns | 0.108ns | |
Td30 | 0.223ns | 0.392ns | 0.019 * | 0.066ns | 0.450ns | 0.036 * | 0.536ns | 0.567ns | 0.486ns | 0.632ns | |
Fi | di | Vb | Fi | di | Vb | |||
|---|---|---|---|---|---|---|---|---|
Plots | n | Min.-Max. (Mean±se) | Min.-Max. (Mean±se) | Min.-Max. (Mean±se) | n | Min.-Max. (Mean±se) | Min.-Max. (Mean±se) | Min.-Max. (Mean±se) |
A. Amegilla calens in the pooled years | B. A. calens in 2021 | |||||||
Untreated | 83 | 1-2 (1±0) | 5.0-33.0 (9.5±0.7) | 2-24 (11±1) | 49 | 1-2 (1±0) | 5.0-33.0 (9.2±0.9) | 2-24 (11±1) |
Parastar | 31 | 1-2 (1±0) | 3.0-60.0 (21.1±2.9) | 1-20 (6±1) | 19 | 1-2 (1±0) | 3.0-60.0 (22.5±4.1) | 1-20 (6±1) |
Cp 10 | 61 | 1-3 (1±0) | 3.0-60.0 (21.0±1.8) | 1-20 (6±0) | 38 | 1-3 (1±0) | 3.0-60.0 (21.7±2.5) | 1-20 (6±1) |
Cp 20 | 58 | 1-2 (1±0) | 5.0-25.0 (11.1±0.8) | 3-24 (10±1) | 37 | 1-2 (1±0) | 5.0-25.0 (11.1±1.0) | 3-24 (10±1) |
Cp 30 | 25 | 1-2 (1±0) | 5.0-8.0 (7.0±0.2) | 8-24 (13±1) | 16 | 1-2 (1±0) | 5.0-8.0 (7.1±0.3) | 8-24 (12±1) |
Ec 10 | 73 | 1-3 (1±0) | 3.0-60.0 (23.6±1.9) | 1-20 (6±0) | 44 | 1-3 (1±0) | 3.0-60.0 (24.2±2.5) | 1-20 (6±1) |
Ec 20 | 67 | 1-2 (1±0) | 3.0-60.0 (23.7±1.9) | 1-20 (5±0) | 41 | 1-2 (1±0) | 3.0-60.0 (23.6±2.6) | 1-20 (5±1) |
Ec 30 | 39 | 1-2 (1±0) | 9.0-50.0 (21.5±1.6) | 1-13 (5±1) | 23 | 1-2 (1±0) | 9.0-50.0 (21.2±2.0) | 1-13 (5±1) |
Td 10 | 49 | 1-2 (1±0) | 5.0-33.0 (13.8±1.0) | 2-24 (8±1) | 28 | 1-2 (1±0) | 5.0-33.0 (13.9±1.3) | 2-24 (8±1) |
Td 20 | 50 | 1-2 (1±0) | 5.0-25.0 (10.7±0.8) | 3-24 (10±1) | 31 | 1-2 (1±0) | 5.0-25.0 (11.7±1.1) | 3-24 (9±1) |
Td 30 | 27 | 1-2 (1±0) | 5.0-8.0 (7.3±0.2) | 8-24 (12±1) | 17 | 1-2 (1±0) | 5.0-8.0 (7.3±0.3 | 8-24 (12±1) |
ANOVA | F(10; 552)=17.939, p<0.001 * | F(10; 332)=10.383, p<0.001 * | ||||||
C. Amegilla sp. in 2021 | D. A. calens in 2022 | |||||||
Untreated | 32 | 1-2 (1±0) | 5.0-33.0 (10.2±1.3) | 2-24 (11±1) | 34 | 1-2 (1±0) | 5.0-33.0 (9.9±1.3) | 2-24 (11±1) |
Parastar | 12 | 1-2 (1±0) | 3.0-60.0 (25.7±6.2) | 1-20 (6±2) | 12 | 1-2 (1±0) | 7.0-56.0 (18.9±4.0) | 1-9 (6±1) |
Cp 10 | 24 | 1-3 (1±0) | 5.0-60.0 (22.6±3.6) | 1-12 (5±1) | 23 | 1-2 (1±0) | 5.0-60.0 (19.8±2.7) | 1-12 (6±1) |
Cp 20 | 24 | 1-2 (1±0) | 5.0-25.0 (10.3±1.1) | 3-24 (10±1) | 21 | 1-2 (1±0) | 5.0-25.0 (11.1±1.4) | 3-24 (10±1) |
Cp 30 | 9 | 1-2 (1±0) | 5.0-8.0 (7.1±0.4) | 8-17 (11±1) | 9 | 1-2 (1±0) | 5.0-8.0 (6.9±0.4) | 8-24 (13±2) |
Ec 10 | 27 | 1-3 (2±0) | 3.0-56.0 (25.8±3.3) | 1-20 (6±1) | 29 | 1-3 (2±0) | 3.0-60.0 (22.8±3.1) | 1-20 (6±1) |
Ec 20 | 24 | 1-2 (1±0) | 3.0-56.0 (26.0±3.3) | 1-20 (5±1) | 26 | 1-2 (1±0) | 5.0-60.0 (23.8±2.9) | 1-13 (5±1) |
Ec 30 | 15 | 1-2 (1±0) | 10.0-33.0 (20.7±2.0) | 2-12 (5±1) | 16 | 1-2 (1±0) | 9.0-50.0 (21.9±2.7) | 1-13 (5±1) |
Td 10 | 18 | 1-2 (1±0) | 6.0-25.0 (14.2±1.4) | 3-15 (7±1) | 21 | 1-2 (1±0) | 5.0-33.0 (13.6±1.7) | 2-24 (9±1) |
Td 20 | 21 | 1-2 (1±0) | 5.0-25.0 (10.6±1.3) | 3-24 (10±1) | 19 | 1-2 (1±0) | 5.0-25.0 (9.1±1.1) | 4-24 (11±1) |
Td 30 | 9 | 1-2 (1±0) | 6.0-8.0 (7.4±0.2) | 8-17 (12±1) | 10 | 1-2 (1±0) | 5.0-8.0 (7.3±0.4) | 8-24 (12±2) |
ANOVA | F(10; 204)=6.181, p<0.001 * | F(10; 209)=7.579, p<0.001 * | ||||||
Aqueous leaves extracts: α’ (p-value) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Unt. | Para | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
A. Pooled campaigns: pooled Amegilla (upper part matrix); Pooled campaigns: Amegilla calens (lower part matrix) | |||||||||||
Unt. | 2x10-5* | 3x10-5* | 0.123ns | 0.446ns | 2x10-5* | 3x10-5* | 1x10-5* | 3x10-5* | 0.084ns | 0.419ns | |
Parastar | 2x10-5* | 0.875ns | 5x10-5* | 3x10-5* | 0.695ns | 0.848ns | 0.893ns | 0.036* | 7x10-5* | 2x10-5* | |
Cp10 | 3x10-5* | 0.983ns | 2x10-5* | 1x10-5* | 0.888ns | 0.698ns | 0.852ns | 0.011* | 2x10-5* | 3x10-5* | |
Cp20 | 0.094ns | 7x10-4* | 5x10-5* | 0.080ns | <0.001* | 2x10-5* | 3x10-5* | 0.011* | 0.893ns | 0.127ns | |
Cp30 | 0.436ns | 3x10-5* | 1x10-5* | 0.060ns | 3x10-5* | 1x10-5* | 1x10-5* | 1x10-3* | 0.080ns | 0.731ns | |
Ec10 | 2x10-5* | 0.885ns | 0.966ns | 2x10-5* | 3x10-5* | 0.848ns | 0.905ns | 0.007* | 2x10-5* | 3x10-5* | |
Ec20 | 3x10-5* | 0.953ns | 0.670ns | 2x10-5* | 1x10-5* | 0.876ns | 0.838ns | 0.005* | 3x10-5* | 1x10-5* | |
Ec30 | 1x10-5* | 0.966ns | 0.832ns | 7x10-5* | 1x10-5* | 0.921ns | 0.831ns | 0.014* | 3x10-5* | 1x10-5* | |
Td10 | 0.002* | 0.019* | 0.014* | 0.148ns | 0.003* | 0.006* | 0.005* | 0.016* | 0.056* | 3x10-4* | |
Td20 | 0.106ns | 7x10-4* | 4x10-5* | 0.733ns | 0.092ns | 2x10-5* | 3x10-5* | 6x10-5* | 0.196ns | 0.105ns | |
Td30 | 0.522ns | 3x10-5* | 3x10-5* | 0.142ns | 0.617ns | 3x10-5* | 1x10-5* | 1x10-5* | 0.010* | 0.167ns | |
B. 2021 campaign: A. calens (upper part matrix) and Amegilla sp. (lower part matrix) | |||||||||||
Unt. | 2x10-4* | 3x10-5* | 0.079ns | 0.698ns | 2x10-5* | 3x10-5* | 1x10-5* | 0.014* | 0.088ns | 0.657ns | |
Parastar | 0.044* | 0.958ns | 0.032* | 9x10-4* | 0.783ns | 0.979ns | 0.978ns | 0.138ns | 0.055ns | 0.002* | |
Cp10 | 3x10-4* | 0.801ns | 0.003* | 4x10-5* | 0.990ns | 0.886ns | 0.922ns | 0.158ns | 0.017* | 1x10-4* | |
Cp20 | 0.707ns | 0.083ns | 0.003* | 0.167ns | 0.002* | 0.002* | 0.008* | 0.385ns | 0.680ns | 0.193ns | |
Cp30 | 0.773ns | 0.164ns | 0.025* | 0.853ns | 5x10-5* | 3x10-5* | 9x10-5* | 0.031* | 0.136ns | 0.757ns | |
Ec10 | 3x10-4* | 0.618ns | 0.853ns | 0.003* | 0.025* | 0.986ns | 0.978ns | 0.082ns | 0.008* | 7x10-5* | |
Ec20 | 3x10-4* | 0.903ns | 0.952ns | 0.003* | 0.027* | 0.967ns | 0.791ns | 0.161ns | 0.013* | 6x10-5* | |
Ec30 | 0.004* | 0.965ns | 0.995ns | 0.020* | 0.062ns | 0.994ns | 0.966ns | 0.235ns | 0.033* | 2x10-4* | |
Td10 | 0.017* | 0.853ns | 0.782ns | 0.782ns | 0.115ns | 0.706ns | 0.856ns | 0.939ns | 0.378ns | 0.050ns | |
Td20 | 0.789ns | 0.092ns | 0.009* | 0.036* | 0.883ns | 0.008* | 0.011* | 0.044* | 0.033* | 0.182ns | |
Td30 | 0.885ns | 0.147ns | 0.017* | 0.891ns | 0.883ns | 0.018* | 0.018* | 0.044* | 0.105ns | 0.894ns | |
Aqueous leaves extracts: α’ (p-value) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Unt. | Para | Cp10 | Cp20 | Cp30 | Ec10 | Ec20 | Ec30 | Td10 | Td20 | Td30 | |
C. A. calens in 2022 | |||||||||||
Unt. | 0.006* | 2x10-4* | 0.272ns | 0.779ns | 4x10-5* | 0.006* | 5x10-4* | 0.142ns | 0.933ns | 0.891ns | |
Parastar | 0.944ns | 0.108ns | 0.014* | 0.993ns | 0.738ns | 0.949ns | 0.263ns | 0.018* | 0.035* | ||
Cp10 | 0.024* | 0.003* | 0.963ns | 0.874ns | 0.972ns | 0.089ns | 0.002* | 0.009* | |||
Cp20 | 0.461ns | 0.009* | 0.009* | 0.043* | 0.478ns | 0.552ns | 0.612ns | ||||
Cp30 | 0.002* | 8x10-4* | 0.003* | 0.234ns | 0.691ns | 0.679ns | |||||
Ec10 | 0.941ns | 0.987ns | 0.030* | 7x10-4* | 0.005* | ||||||
Ec20 | 0.997 ns | 0.063 ns | 3x10-4 * | 0.003 * | |||||||
Ec30 | 0.178 ns | 0.003 * | 0.010 * | ||||||||
Td10 | 0.308 ns | 0.378 ns | |||||||||
Td20 | 0.719 ns | ||||||||||
α | Significance Level |
α’ | Bonferroni Corrected Significance Level |
A. calens | Amegilla calens Le Peletier. 1841 |
A1,000 | Abundances Per 1,000 Flowers |
ANOVA | Analysis of Variance |
Ap. mellifera | Apis mellifera Linnaeus, 1758 |
IRD | Institut de Recherche Agricole Pour le Développement / Agricultural Research Institute for Development |
Ca. procera | Calotropis procera (Aiton) Aiton, 1811 |
Cp10 | 10% Leaves Extract of Ca. procera |
Cp20 | 20% Extract of Ca. procera |
Cp30 | 30% Extract of Ca. procera |
di | Delay of the Visits |
E. camaldulensis | Eucalyptus camaldulensis Dehnh., 1832 |
Ec10 | 10% Extract of E. camaldulensis |
Ec20 | 20% Extract of E. camaldulensis |
Ec30 | 30% Extract of E. camaldulensis |
Fi | Visited Flowers |
Max | Maximum |
Min. | Minimum |
MINADER | Ministry of Agriculture and Rural Development (Cameroon) |
ns | Not Significant |
p-value | Probability Value |
Para. | Parastar |
se | Standard Error |
Ti. diversifolia | Tithonia diversifolia (Hemsley) Gray, 1883 |
Td10 | 10% Extract of Ti. diversifolia |
Td20 | 20% Extract of Ti. diversifolia |
Td30 | 30% Extract of Ti. diversifolia |
Unt. | Untreated |
V. unguiculata | Vigna unguiculata (L.) Walp., 1843 |
Vb | Foraging Speed |
| [1] | Ige, O. E., Olotuah, O. F., Akerele, V.. Floral biology and pollination ecology of cowpea (Vigna unguiculata L. Walp). Modern Applied Science. 2011, 5(4), 74-82. |
| [2] | Otiobo Atibita, E. N. A., Tchuenguem Fohouo, F.-N., Djieto-Lordon, C. Activité de butinage et de pollinisation de Apis mellifera adansonii Latreille (Hymenoptera: Apidae) sur les fleurs d’Oxalis barrelieri (Oxalidaceae) à Yaoundé (Cameroun). Entomologie Faunistique. 2015, 68, 101-108. |
| [3] | Taimanga, Tchuenguem Fohouo, F.-N. Pollination efficiency of Apis mellifera Linnaeus 1758 (Hymenoptera: Apidae) on Mimosa pudica Linnaeus 1753 (Fabaceae) inflorescences at Yassa (Douala - Cameroon). Journal of Entomology and Zoology Studies. 2018, 6(5), 2027-2033. |
| [4] | Zra, G. V., Mazi, S., Tchuenguem Fohouo, F.-N. Pollination efficiency of Dactylurina staudingeri (Hymenoptera: Apidae) on Psorospermum febrifugum (Hypericaceae) at dang (Ngaoundere, Cameroon). Journal of Entomology and Zoology Studies. 2020, 8(1), 216-224. |
| [5] | Adamou, M., Kosini, D., Tchoubou-Salé, A., Massah, O. D., Tchocgnia, T. F. C., Mohammadou, M., Youssoufa, O., Nukenine, E. N. Impact of aqueous extracts of Cassia occidentalis, Eucalyptus camaldulensis and Hyptis suaveolens on the entomofauna and the seed yield of Gossypium hirsutum at Bokle (Garoua, Cameroon). Heliyon. 2022, 8(10), e 10937. |
| [6] | Mweke, A., Akutse, K. S., Ulrichs, C., Fiaboe, K. K. M., Maniania, N. K., Ekesi, S. Integrated management of Aphis craccivora in cowpea using intercropping and entomopathogenic fungi under field conditions. Journal of Fungi. 2020, 6(2), 1-16. |
| [7] | Alam, M. Z., Crump, A. R., Haque, M. M., Islam, M. S., Hossain, E., Hasan, S. B., Hasan, S. B., Hossain, M. S. Effects of Integrated Pest Management on Pest Damage and Yield Components in a Rice Agro-Ecosystem in the Barisal Region of Bangladesh. Frontiers in Environmental Science. 2016, 4, 22. |
| [8] | Mohammadou, M., Adamou, M., Taïmanga, Kosini, D., Kenne, M. Seed Yield Improvement in Vigna unguiculata (L.) (Fabaceae): Efficiency of Pollinators and Impact of Aqueous Leaf Extract of Three Plant Species in North Cameroon. Asian Journal of Crop Science. 2023a, 8(3), 146-172. |
| [9] | Mohammadou, M., Fouelifack-Nintidem, B., Adamou, M., Taimanga, Kossini, D., Tsekane, S. J., Ngamaleu-Siewe, B., Kenne, E. L., Yomon, A. K., Kenne, M. Diversity and Abundance of Pest Insects Associated with Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae) in Bockle and Dang Localities (North-Cameroon). American Journal of Entomology. 2023b, 7(2), 38-61. |
| [10] | Ngegba, P. M., Cui, G., Khalid, M. Z., Zhong, G. Use of Botanical Pesticides in Agriculture as an Alternative to Synthetic Pesticides. Agriculture. 2022, 12(5), 600. |
| [11] | Nghia, N. T., Srivastava, P. Biodiversity of beneficial insect associated with cowpea at Pantnagar, Uttarakhand, India. OMONRICE. 2015, 20, 73-79. |
| [12] | Anusha, C., Natikar, P. K., Balikai, R. A. Insect pests of cowpea and their management – A review. Journal of Experimental Zoology, India. 2016, 19(2), 635-642. |
| [13] | Zaki, A. Y., Aly, A. I. Biodiversity of Spider and Other Arthropods Inhabiting Cowpea under Effect of Fish Culture Water and Nitrogen Fertilization and its effect on Yield and Protein at Fayoum Governorate, Egypt. ACARINES. 2018, 12, 87-98. |
| [14] | Manap, T., Fajri, M. Diversity of Bees and Wasp (Hymenoptera) in Cowpea (Vigna sinensis L.) in Agricultural Area at Martapura District, Banjar Regency, South Kalimantan. Journal of Science and Technology. 2020, 9, 29-33. |
| [15] | Apriyani, S., Kurnia, A., Sutriadi, M. T. Diversity of insect on cowpea cropping in rain fed land. IOP Conference Series: Earth and Environmental Science. 2021, 739, 012068. |
| [16] |
Ministry of Agriculture and Rural Development (Cameroon). MINADER: Liste des pesticides homologués au Cameroun au 18 Avril 2019. Liste reservée au grand public. Ministère de l’Agriculture et de Développement Rural. Commission Nationale d’Homologation des Produits Phytosanitaires et de Certification des Appareils de Traitement (CNHPPCZT), Yaoundé, Cameroun, 2019. Available from:
https://drcq-minader.org/docs/Liste_Pesticides_Homologues_042019.pdf |
| [17] | Nantia, E. A., Manfo, T. F. P., Sonchieu, J., Choumessi, T. A., Bopuwouo, R. H., Kakwang, F. I. Parastar insecticide induced changes in reproductive parameters and testicular oxidative stress biomarkers in wistar male rats from Santa (Cameroon). Toxicology and Industrial Health. 2018, 34(7), 499-506. |
| [18] | Kumari, K. A., Kumar, K. N. R., Rao, C. N. Adverse effects of chemical fertilizers and pesticides on human health and environment. Journal of Chemical and Pharmaceutical Research. 2014, 3, 150-151. |
| [19] | Sonchieu, J., Ngassoum, M. B., Nantia, Akono, E., Laxman, P. S. Pesticide Applications on Some Vegetables Cultivated and Health Implications in Santa, North-West Cameroon. SSRG. International Journal of Agriculture and Environmental Research. 2018, 4(2), 39-46. |
| [20] | Siyunda, A. C., Mwila, N., Mwala, M., Munyinda, K. L., Kamfwa, K., Kamfwa, K., Chipabika, G. Nshimbi, D. Laboratory Screening of Cowpea (Vigna unguiculata) Genotypes against Pulse Beetle, Callosobruchus maculates. International Journal of Business and Social Science. 2022, 6(1), 85-93. |
| [21] | Sreekanth. Field evaluation of certain leaf extracts for the control of mussel scale (Lepidosaphes piperis Gr.) in Black pepper (Piper nigrum L.). Journal of Biopesticides. 2013, 6(1), 1-5. |
| [22] | Barry, B. R., Ngakou, A. T. M., Nukenine, N. E. The incidence of aqueous neem leaves (Azadirachta indica A. Juss) extracts and Matarhizium anisopliae Mech on crowpea flowers Thrips (Megolurothips sjostedti) and Field in Ngaoundere. Journal of Entomology and Zoology Studies. 2019, 7(5), 333-338. |
| [23] | Moudelsia, O. B., Otiobo Atibita, N. E., Heumou, C. R., Pando, J. B., Djonwangwe, D. Comportement de Butinage de Trois Abeilles Sauvages (Amegilla sp., Sphecodes albilabris et Lasioglossum albipes) sur les Fleurs Mâles de Zea mays (Poaceae) à Maroua, Cameroun. European Scientific Journal, ESJ. 2022, 18(27), 12. |
| [24] | Mazi, S., Kingha, T. B. M., Adamou, M., Yatahaï, C. M. Impact of the foraging activity of Apis mellifera (Hymenoptera: Apidae) on increasing yields of Gossypium hirsutum (Malvaceae) in Djoumassi (Garoua, Cameroon). GSC Biological and Pharmaceutical Sciences. 2020, 12(1), 255-266. |
| [25] |
Integrated Taxonomic Information System. ITIS: Amegilla Friese, 1897. Taxonomic Serial No.: 634005 on-line database. Available from: www. itis. gov, CC0
https://doi.org/10.5066/F7KH0KBK (accessed 12 May 2024) |
| [26] | Statistical Package for the Social Sciences. SPSS: SigmaStat for Windows version 2. 03. IBM SPSS Inc., Chicago, IL; 1992-1997. |
| [27] | Rice, W. Analyzing tables of statistical tests. Evolution, 1989, 43(1), 223-225. |
| [28] | Issaya, I., Kengni, B., Tope, S., Tchuenguem Fohouo, F. Pollination efficiency of Amegilla calens (Hymenoptera: Apidae) on Gossypium hirsutum L. (Malvaceae) variety L457 flowers at Meskine (Maroua, Cameroon). Agricultural Sciences. 2021, 12, 1150-1167. |
| [29] | Wang, X., Liu, H., Li, X., Song, Y., Li, C., Jin, L. Correlations between environmental factors and wild bee behavior on alfalfa (Medicago sativa) in Northwestern China. Environmental Entomology. 2009, 38, 1480–1484. |
| [30] | Malovrh, K., Ravnjak, B., Bavcon, J., Križman, M. Nectar production and three main sugars in nectar of Salvia pratensis and Salvia glutinosa in correlation with abiotic factors. Agriculture. 2024, 14, 668. |
| [31] | Manggoel, W., Uguru, M. I. Evidence of maternal effect on the inheritance of flowering time in cowpea (Vigna unguiculata (L.) Walp.). International Journal of Plant Breeding and Genetics. 2012, 6, 1-16. |
| [32] | Dubey, N. K. Shukla, R., Kumar, A., Singh, P., Prakash, B. Prospects of botanical pesticides in sustainable agriculture. Current Science. 2020, 98(4), 479-480. |
| [33] | Kot, I., Lisecka, M., Kmieć, K., Golan, K., Górska-Drabik, E., Kiljanek, T., Zimowska, B., Skwaryło-Bednarz, B. Visitation of Apis mellifera L. in runner bean (Phaseolus coccineus L.) and its exposure to seasonal agrochemicals in agroecosystems. Agriculture. 2023, 13, 2138. |
| [34] | Lin, Y.-C., Lu, Y.-H., Tang, C.-K., Yang, E.-C., Wu. Y.-L. Honey bee foraging ability suppressed by imidacloprid can be ameliorated by adding adenosine. Environment and Pollution. 2023, 332, 121920. |
| [35] | Raymann, K., Motta, E. V. S., Girard, C., Riddington, I. M., Dinser, J. A., Moran, N. A. Imidacloprid decreases honey bee survival rates but does not affect the gut microbiome. Applied Environmental Microbiology. 2018, 84, e00545-18. |
| [36] | Feng, B., Qian, K., Du, Y.-J. Floral Volatiles from Vigna unguiculata Are Olfactory and Gustatory Stimulants for Oviposition by the Bean Pod Borer Moth Maruca vitrata. Insects. 2017, 8(2), 60. |
| [37] | Pharaon, M. A., Douka, C., Dounia, Eloundou, C. E., Tchuenguem Fohouo, F.-N. Pollination efficiency of Apis mellifera L. (Hymenoptera: Apidae) on flowers of Vigna unguiculata (L.) Walp. (Fabaceae) at Bilone (Obala, Cameroon). International Journal of Bioscience. 2019, 14(1), 1-11. |
| [38] | Anderson, N. L., Harmon-Threatt, A. N. Chronic contact with realistic soil concentrations of imidacloprid affects the mass, immature development speed, and adult longevity of solitary bees. Scientific Reports. 2019, 9, 3724. |
| [39] |
Gervais, J. A., Luukinen, B., Buhl, K., Stone, D. Imidacloprid General Fact Sheet. National Pesticide Information Center (NPIC), Oregon State University Extension Services, 2010. Available from:
http://npic.orst.edu/factsheets/imidagen.html (accessed 19 April 2022) |
| [40] | Bonmatin, J. M., Moineau, I., Charvet, R., Colin, M. E., Fleche, C., Bengach, E. R. Behaviour of imidacloprid in fields. Toxicity for honey bees. In Environmental Chemistry. Lichtfouse, E., Schwarzbauer, J., Robert, D., Eds., Springer, Berlin Heidelberg, 2005, pp. 483–494. |
| [41] | Sharma, S., Singh, B. Metabolism and persistence of imidacloprid in different types of soils under laboratory conditions. International Journal of Environmental Analytical Chemistry. 2014, 94, 1100-1112. |
| [42] | Yang, E., Chuang, Y., Chen, Y., Chang, L. Abnormal foraging behavior induced by sublethal dosage of imidacloprid in the honey bee (Hymenoptera: Apidae). Journal of Economic Entomology. 2008, 101, 1743-1748. |
APA Style
Taimanga, Adamou, M., Tchindebe, G., Mohammadou, M., Youssoufa, O., et al. (2024). Effect of the Botanical Insecticides on Amegilla Friese, 1897 (Hymenoptera: Apidae) Foraging on the Cowpea Flowers in Dang (Adamaoua, North-Cameroon). American Journal of Entomology, 8(3), 76-101. https://doi.org/10.11648/j.aje.20240803.13
ACS Style
Taimanga; Adamou, M.; Tchindebe, G.; Mohammadou, M.; Youssoufa, O., et al. Effect of the Botanical Insecticides on Amegilla Friese, 1897 (Hymenoptera: Apidae) Foraging on the Cowpea Flowers in Dang (Adamaoua, North-Cameroon). Am. J. Entomol. 2024, 8(3), 76-101. doi: 10.11648/j.aje.20240803.13
AMA Style
Taimanga, Adamou M, Tchindebe G, Mohammadou M, Youssoufa O, et al. Effect of the Botanical Insecticides on Amegilla Friese, 1897 (Hymenoptera: Apidae) Foraging on the Cowpea Flowers in Dang (Adamaoua, North-Cameroon). Am J Entomol. 2024;8(3):76-101. doi: 10.11648/j.aje.20240803.13
@article{10.11648/j.aje.20240803.13,
author = {Taimanga and Moise Adamou and Georges Tchindebe and Moukhtar Mohammadou and Ousmana Youssoufa and Boris Fouelifack-Nintidem and Alice Virginie Tchiaze Ifoue and Andrea Sarah Kenne Toukem and Odette Massah Dabole and Oumarou Abdoul Aziz and Abraham Tchoubou-Sale and Sedrick Junior Tsekane and Daniel Kosini and Pharaon Auguste Mbianda and Martin Kenne},
title = {Effect of the Botanical Insecticides on Amegilla Friese, 1897 (Hymenoptera: Apidae) Foraging on the Cowpea Flowers in Dang (Adamaoua, North-Cameroon)
},
journal = {American Journal of Entomology},
volume = {8},
number = {3},
pages = {76-101},
doi = {10.11648/j.aje.20240803.13},
url = {https://doi.org/10.11648/j.aje.20240803.13},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.aje.20240803.13},
abstract = {Synthetic pesticides present risks of pollution of the environment, humans and livestock and the alternative proposed today is to use botanical extracts in the fields against crop pests. But in North Cameroon, little information exists concerning the effect of these extracts on useful pollinating insects in general and no information exists in particular on foragers of the genus Amegilla Friese, 1897 (Apidae: Apinae: Anthophorini). The frequency and foraging activities of Amegilla, on newly blooming flowers of Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae) were recorded during five consecutive days in 2021 and 2022 planting campaigns. Plants were divided into untreated plots and plots treated using the synthetic insecticide Parastar (l p.c..ha-1) or 10%, 20% and 30% aqueous leaf extracts of Calotropis procera (Aiton) Aiton, 1811 (Gentianales: Apocynaceae), Eucalyptus camaldulensis Dehnh., 1832 (Myrtales: Myrtaceae) and Tithonia diversifolia (Hemsley) Gray, 1883 (Asterales: Asteraceae) respectively. Among 8,987 insects collected (48.9% in 2021), Amegila calens Le Peletier. 1841 with stockier foragers (2021 campaign: 2.2% of the total collection, entomophily FA. calens=4.5%; 2022 campaign: 0.7%, FA. calens=1.3%; pooled campaigns: 2.9%, FA. calens=2.9%) and Amegilla sp. with slender foragers (2021: 3.8%, FAmegilla sp.=7.7%; 2022: no data) were recorded. Foragers started activity from 6 a.m. and stopped foraging before noon, with a peak of activity in 8 to 9 a.m. time slot for A. calens and 10 to 11 a.m. time slot for Amegilla sp.. During the five consecutive days from the first blooming day of the flowers, 598 visits (89.8% in 2021 and 10.2% in 2022) were recorded with a peak of visits during the 3rd day and then declined until it stopped during the 5th day. Treatments including the synthetic insecticide (which was the most repellent to the wild bees), did not significantly reduce the frequency of visits. But 20% aqueous extract of Ca. procera showed a significant increased of the mean duration of visits of the bees, compare to the results recorded in Parastar-treated plots. Therefore, the tested extracts, especially 20% aqueous leaves extract of Ca. procera may be recommended to control field insect pests and for preservation of foraging activities of Amegilla genus.
},
year = {2024}
}
TY - JOUR T1 - Effect of the Botanical Insecticides on Amegilla Friese, 1897 (Hymenoptera: Apidae) Foraging on the Cowpea Flowers in Dang (Adamaoua, North-Cameroon) AU - Taimanga AU - Moise Adamou AU - Georges Tchindebe AU - Moukhtar Mohammadou AU - Ousmana Youssoufa AU - Boris Fouelifack-Nintidem AU - Alice Virginie Tchiaze Ifoue AU - Andrea Sarah Kenne Toukem AU - Odette Massah Dabole AU - Oumarou Abdoul Aziz AU - Abraham Tchoubou-Sale AU - Sedrick Junior Tsekane AU - Daniel Kosini AU - Pharaon Auguste Mbianda AU - Martin Kenne Y1 - 2024/08/27 PY - 2024 N1 - https://doi.org/10.11648/j.aje.20240803.13 DO - 10.11648/j.aje.20240803.13 T2 - American Journal of Entomology JF - American Journal of Entomology JO - American Journal of Entomology SP - 76 EP - 101 PB - Science Publishing Group SN - 2640-0537 UR - https://doi.org/10.11648/j.aje.20240803.13 AB - Synthetic pesticides present risks of pollution of the environment, humans and livestock and the alternative proposed today is to use botanical extracts in the fields against crop pests. But in North Cameroon, little information exists concerning the effect of these extracts on useful pollinating insects in general and no information exists in particular on foragers of the genus Amegilla Friese, 1897 (Apidae: Apinae: Anthophorini). The frequency and foraging activities of Amegilla, on newly blooming flowers of Vigna unguiculata (L.) Walp., 1843 (Fabales: Fabaceae) were recorded during five consecutive days in 2021 and 2022 planting campaigns. Plants were divided into untreated plots and plots treated using the synthetic insecticide Parastar (l p.c..ha-1) or 10%, 20% and 30% aqueous leaf extracts of Calotropis procera (Aiton) Aiton, 1811 (Gentianales: Apocynaceae), Eucalyptus camaldulensis Dehnh., 1832 (Myrtales: Myrtaceae) and Tithonia diversifolia (Hemsley) Gray, 1883 (Asterales: Asteraceae) respectively. Among 8,987 insects collected (48.9% in 2021), Amegila calens Le Peletier. 1841 with stockier foragers (2021 campaign: 2.2% of the total collection, entomophily FA. calens=4.5%; 2022 campaign: 0.7%, FA. calens=1.3%; pooled campaigns: 2.9%, FA. calens=2.9%) and Amegilla sp. with slender foragers (2021: 3.8%, FAmegilla sp.=7.7%; 2022: no data) were recorded. Foragers started activity from 6 a.m. and stopped foraging before noon, with a peak of activity in 8 to 9 a.m. time slot for A. calens and 10 to 11 a.m. time slot for Amegilla sp.. During the five consecutive days from the first blooming day of the flowers, 598 visits (89.8% in 2021 and 10.2% in 2022) were recorded with a peak of visits during the 3rd day and then declined until it stopped during the 5th day. Treatments including the synthetic insecticide (which was the most repellent to the wild bees), did not significantly reduce the frequency of visits. But 20% aqueous extract of Ca. procera showed a significant increased of the mean duration of visits of the bees, compare to the results recorded in Parastar-treated plots. Therefore, the tested extracts, especially 20% aqueous leaves extract of Ca. procera may be recommended to control field insect pests and for preservation of foraging activities of Amegilla genus. VL - 8 IS - 3 ER -
Department of Agronomy, Institute of Fisheries and Aquatic Sciences, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cotton (Gossypium hirsutum), Pollinization
Laboratory of Applied Zoology, Faculty of Sciences, University of Ngaoundere, Ngaoundere, Cameroon; Faculty of Medicine and Biomedical Sciences of Garoua, University of Garoua, Garoua, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cotton (Gossypium hirsutum), Pollinization
Department of Agronomy, Institute of Fisheries and Aquatic Sciences, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cotton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cotton (Gossypium hirsutum), Pollinization
Laboratory of Applied Zoology, Faculty of Sciences, University of Ngaoundere, Ngaoundere, Cameroon; Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cotton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Pest control, Entomology, Biology of animal populations, Insects ecology, Applied entomology, Animal ethology
Laboratory of Biology and Physiology of Plant Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Plant physiology, Soil fertilization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Toxicology, quality of life, Biostatistics, Animal biology, Public health
Laboratory of Applied Zoology, Faculty of Sciences, University of Ngaoundere, Ngaoundere, Cameroon; Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cotton (Gossypium hirsutum), Pollinization
Laboratory of Applied Zoology, Faculty of Sciences, University of Ngaoundere, Ngaoundere, Cameroon; Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Biopesticides, Insect pest control, Biological agriculture, Bees, Biological agriculture, Pollinization
Faculty of Medicine and Biomedical Sciences of Garoua, University of Garoua, Garoua, Cameroon; Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Botanical insecticides, Bees, Biological agriculture, Yield agricultural products, Cotton (Gossypium hirsutum), Pollinization
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied Zoology, Quality of life and biostatistics, Wildlife Protection, Control of protected areas, Animal Ethology, Animal Ecology
Laboratory of Applied Zoology, Faculty of Sciences, University of Ngaoundere, Ngaoundere, Cameroon; Faculty of Medicine and Biomedical Sciences of Garoua, University of Garoua, Garoua, Cameroon
Research Fields: Conceptualization, Data curation, Investigation, Methodology
Laboratory of Biology and Physiology of Plant Organisms, University of Douala, Douala, Cameroon
Research Fields: Applied entomology, Insects biology, biostatistics and pollinators, Apiculture, Insects-plants interactions, Animal Ethology, Animal Ecology
Laboratory of Biology and Physiology of Animal Organisms, University of Douala, Douala, Cameroon
Research Fields: Biostatistics, Biology of the Animal Populations, Entomology, Myrmecology, Animal Ethology, Animal Ecology, sociobiology, applied entomology, plant protection, Biological control, pest insects
Figure 1. Location of the study site in Dang. A: Adamaoua Region in Cameroon; B: Dang locality (Ngaoundere III suburb).

Figure 2. Illustration of Amagilla Friese, 1897 (Hymenoptera: Apidae) recorded in Dang (North-Cameroon). A: top view of an adult of Amegilla calens Le Peletier, 1841 (Hymenoptera: Apidae); B: top view of an adult of Amegilla sp. Friese, 1897 (Hymenoptera: Apidae).
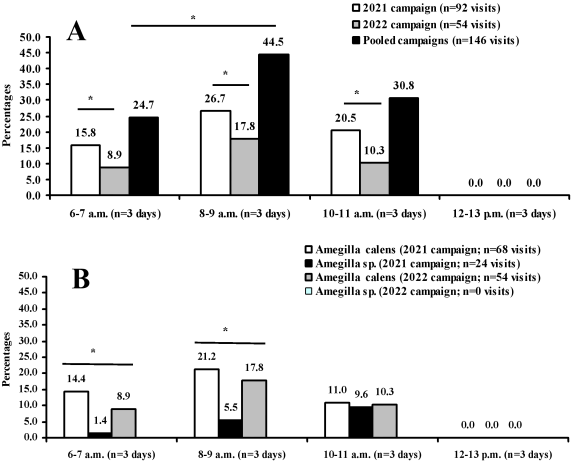
Figure 3. Daily activity rhythm of Amegilla on cowpea flowers at Dang (Adamaoua Region, North-Cameroon) in 2021 and 2022. A: overall daily activity rhythm of the pooled species showing the peak of activity in 8-9 a.m. time slot; B: daily activity rhythm of Amegilla calens and Amegilla sp. foragers during each campaign.
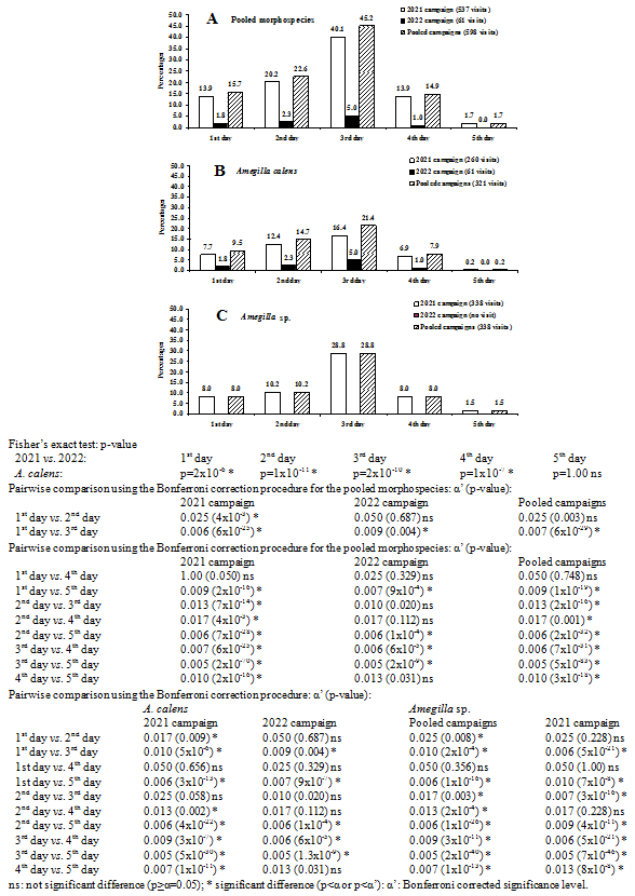
Figure 4. Activity rhythm of Amegilla in five consecutive days from the first day of the flower blooming.
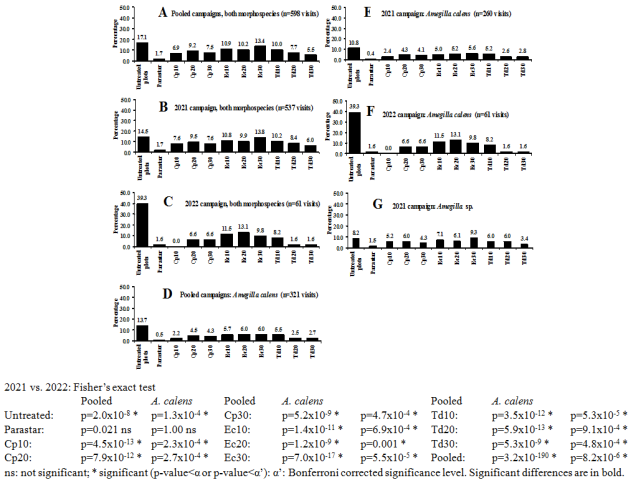
Figure 5. Effect of the chemical treatments on the visiting rate of the cowpea flowers at Dang locality (Adamaoua Region, North-Cameroon) in 2021 and 2022. Parastar: synthetic insecticide (mixture of 20 g.l-1 of imidacloprid and 20 g.l-1 of lamda-cyhalothrin, 1 p.c..ha-1); Cp10, Cp20 and Cp30: 10%, 20% and 30% respectively of aqueous leaves extract of Calotropis procera (Aiton) Aiton, 1811 (Gentianales: Apocynaceae); Ec10, Ec20 and Ec30: 10%, 20% and 30% respectively of Eucalyptus camaldulensis Dehnh., 1832 (Myrtales: Myrtaceae); Td10, Td20 and Td30: 10%, 20% and 30% respectively of the aqueous leaves extract of Tithonia diversifolia (Hemsley) Gray, 1883 (Asterales: Asteraceae).
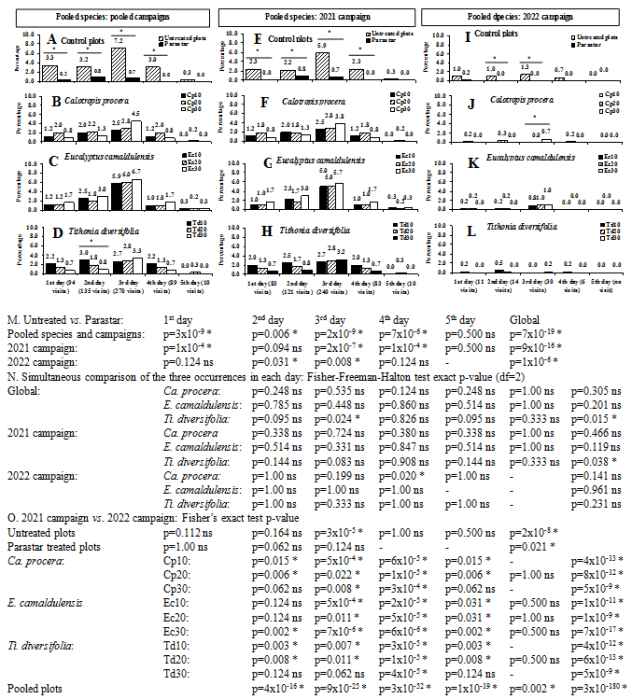
Figure 6. Effect of the chemical treatments during five first consecutive days of flower blooming on the visiting rate of the cowpea flowers at Dang locality (Adamaoua Region, North-Cameroon) during 2021 and 2022 campaigns. ns: not significant difference (p≥0.05); * significant difference (p<0.05). Abbreviations Cp10, Cp20, Cp30, Ec10, Ec20, Ec30, Td10, Td20 and Td30 are presented in Figure 5.
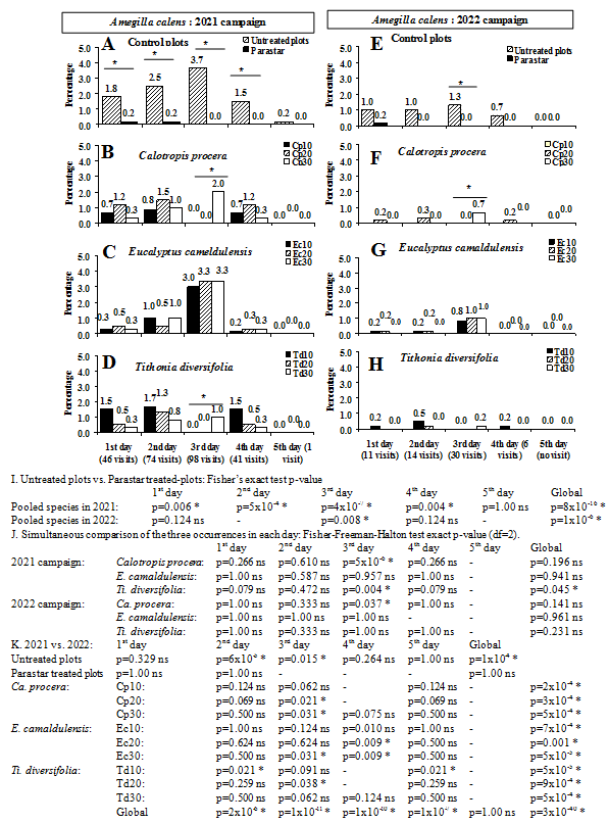
Figure 7. Effect of the chemical treatments during five first consecutive days of flower blooming on the visiting rate of Amegilla calens at Dang (Adamaoua Region, North-Cameroon) in 2021 and 2022. ns: not significant difference (p≥0.05); * significant difference (p<0.05). Cp10, Cp20, Cp30, Ec10, Ec20, Ec30, Td10, Td20 and Td30 are presented in Figure 5.
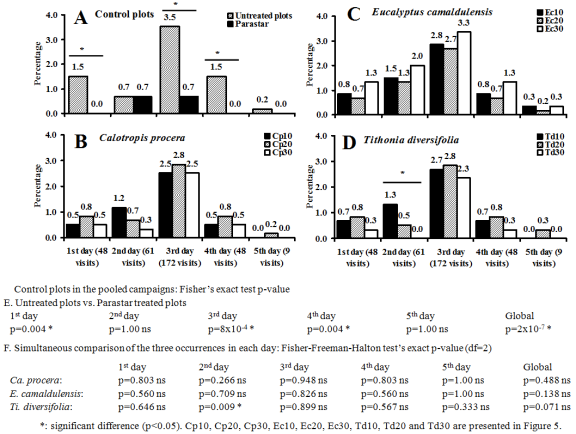
Figure 8. Effect of the chemical treatments during five first consecutive days of flower blooming on the visiting rate of Amegilla sp. on the cowpea flowers at Dang (Adamaoua Region, North-Cameroon) during the 2021 campaign. ns: not significant difference (p≥0.05).
Information

