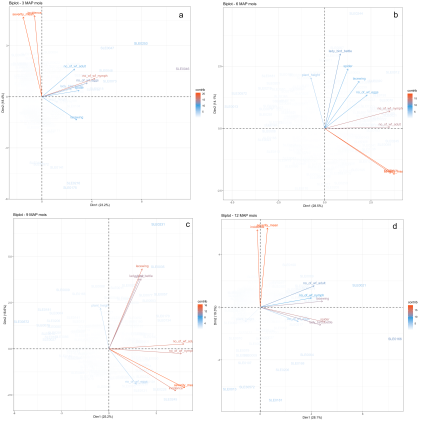
Bemisia tabaci is a major pest of cassava in sub-Saharan Africa, causing yield losses through direct feeding and its role in transmitting cassava mosaic disease (CMD). Natural enemies such as lacewings, ladybird beetles, and spiders provide valuable biological control services, yet their interactions with different whitefly developmental stages and plant structural traits remain insufficiently characterized. This study examined the dynamics among natural enemies, whitefly eggs, nymphs, adults, and plant height across 3, 6, 9, and 12 months after planting (MAP) under field conditions. The trial was conducted under natural cassava production conditions during 2020/2021 cropping season at the upland experimental site of the School of Agriculture and Food Sciences, Njala University. A total of 270 cassava genotypes comprising 268 local varieties and 2 improved checks (SLICASS 4 and SLICASS 6) were laid out in an augmented randomized design with four blocks. Results showed that lacewings and spiders strongly tracked nymph and adult whitefly populations, while ladybird beetles showed weaker associations. Principal Component Analysis (PCA) revealed alignment of predators with pest pressure during mid- and late season, whereas plant height exhibited minimal influence. Findings underscore the central role of lacewings and spiders in early and sustained suppression of whitefly populations, highlighting the importance of conservation-based integrated pest management (IPM) strategies. Findings serve as useful guide for conservation biological control as a primary IPM strategy for the enhancement of habitats for effective predators (lacewings and spiders) of the whitefly through reduced pesticide use, ground vegetation retention, intercropping, and maintenance of natural refuge habitats.
| Published in | American Journal of Entomology (Volume 10, Issue 1) |
| DOI | 10.11648/j.aje.20261001.11 |
| Page(s) | 1-15 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Natural Enemies, Whitefly Developmental Stages, Cassava Mosaic Disease, Prevalence, Correlation, Cassava
Trait | LW | LBB | SP | NWE | NWN | NWA |
|---|---|---|---|---|---|---|
Correlation coefficient at 3 MAP | ||||||
LW | 1.00 | |||||
LBB | 0.06 | 1.00 | ||||
SP | 0.22*** | 0.16* | 1.00 | |||
NEW | 0.09 | 0.09 | 0.15* | 1.00 | ||
NWN | 0.21*** | 0.19** | 0.23*** | 0.63*** | 1.00 | |
NWA | 0.13* | 0.15* | 0.23*** | 0.13* | 0.22*** | 1.00 |
Correlation coefficient at 6 MAP | ||||||
LW | 1.00 | |||||
LBB | 0.17*** | 1.00 | ||||
SP | 0.12* | 0.17*** | 1.00 | |||
NEW | 0.10 | 0.07 | 0.14* | 1.00 | ||
NWN | 0.21*** | 0.11 | 0.10 | 0.40*** | 1.00 | |
NWA | 0.22*** | 0.05 | 0.17*** | 0.22*** | 0.49*** | 1.00 |
Correlation coefficient at 9 MAP | ||||||
LW | 1.00 | |||||
LBB | 0.23 | 1.00 | ||||
SP | 0.22*** | 0.33*** | 1.00 | |||
NEW | -0.07 | -0.15* | 0.08 | 1.00 | ||
NWN | 0.12 | 0.09 | 0.13* | 0.24*** | 1.00 | |
NWA | 0.28*** | 0.08 | 0.07 | 0.13* | 0.59*** | 1.00 |
Correlation coefficient at 12 MAP | ||||||
LW | 1.00 | |||||
LBB | 0.34*** | 1.00 | ||||
SP | 0.43*** | 0.47*** | 1.00 | |||
NEW | 0.16** | 0.42*** | 0.30*** | 1.00 | ||
NWN | 0.35*** | 0.24*** | 0.26*** | 0.03 | 1.00 | |
NWA | 0.42*** | 0.24*** | 0.30*** | -0.001 | 0.42*** | 1.00 |
Variable | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 |
|---|---|---|---|---|---|---|---|---|---|
PHT | 0.227 | 0.055 | 0.719 | 0.107 | 0.483 | 0.024 | 0.418 | 0.007 | 0.093 |
LW | 0.503 | 0.244 | 0.352 | 0.48 | 0.054 | 0.373 | 0.415 | 0.113 | 0.07 |
LBB | 0.428 | 0.084 | 0.231 | 0.786 | 0.06 | 0.325 | 0.151 | 0.017 | 0.078 |
SP | 0.537 | 0.066 | 0.17 | 0.019 | 0.73 | 0.041 | 0.378 | 0.032 | 0.013 |
NEW | 0.654 | 0.145 | 0.528 | 0.094 | 0.289 | 0.005 | 0.223 | 0.189 | 0.308 |
NWN | 0.763 | 0.193 | 0.418 | 0.065 | 0.137 | 0.053 | 0.067 | 0.191 | 0.373 |
NWA | 0.473 | 0.293 | 0.337 | 0.057 | 0.02 | 0.705 | 0.269 | 0.056 | 0.039 |
CMDS | 0.269 | 0.841 | 0.04 | 0.011 | 0.005 | 0.163 | 0.023 | 0.404 | 0.167 |
CMDI | 0.106 | 0.857 | 0.139 | 0.198 | 0.011 | 0.142 | 0.014 | 0.374 | 0.19 |
EV | 2.092 | 1.659 | 1.312 | 0.916 | 0.875 | 0.794 | 0.64 | 0.393 | 0.319 |
PV | 23.25 | 18.44 | 14.58 | 10.18 | 9.72 | 8.82 | 7.11 | 4.36 | 3.54 |
CV | 23.25 | 41.68 | 56.26 | 66.44 | 76.16 | 84.98 | 92.09 | 96.46 | 100 |
Variable | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 |
|---|---|---|---|---|---|---|---|---|---|
PHT | -0.150 | 0.411 | -0.151 | 0.772 | 0.328 | 0.277 | -0.052 | 0.013 | 0.045 |
LW | 0.375 | 0.382 | 0.280 | 0.135 | -0.659 | 0.288 | 0.316 | -0.003 | 0.024 |
LBB | 0.170 | 0.604 | 0.435 | 0.061 | 0.043 | -0.607 | -0.198 | 0.065 | 0.015 |
SP | 0.253 | 0.483 | 0.226 | -0.501 | 0.476 | 0.389 | 0.107 | -0.100 | -0.051 |
NEW | 0.463 | 0.271 | -0.623 | -0.096 | 0.094 | -0.265 | 0.430 | 0.223 | 0.025 |
NWN | 0.710 | 0.141 | -0.386 | 0.018 | -0.132 | -0.071 | -0.255 | -0.487 | -0.046 |
NWA | 0.707 | 0.012 | -0.118 | -0.093 | -0.104 | 0.256 | -0.497 | 0.392 | -0.007 |
CMDS | 0.760 | -0.369 | 0.256 | 0.103 | 0.211 | -0.026 | 0.110 | -0.053 | 0.387 |
CMDI | 0.718 | -0.361 | 0.268 | 0.281 | 0.184 | -0.068 | 0.182 | 0.031 | -0.362 |
EV | 2.567 | 1.273 | 1.030 | 0.977 | 0.886 | 0.825 | 0.695 | 0.459 | 0.289 |
PV | 28.53 | 14.14 | 11.44 | 10.85 | 9.84 | 9.16 | 7.72 | 5.10 | 3.22 |
CV | 28.53 | 42.67 | 54.11 | 64.97 | 74.806 | 83.97 | 91.69 | 96.79 | 100 |
Variable | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 |
|---|---|---|---|---|---|---|---|---|---|
PHT | -0.081 | 0.332 | 0.399 | 0.599 | 0.464 | 0.378 | 0.076 | 0.022 | -0.029 |
LW | 0.317 | 0.653 | 0.131 | 0.33 | -0.06 | -0.565 | 0.016 | -0.131 | 0.08 |
LBB | 0.307 | 0.572 | -0.475 | -0.213 | -0.073 | 0.207 | 0.507 | 0.057 | 0.001 |
SP | 0.289 | 0.571 | -0.112 | -0.461 | 0.391 | 0.082 | -0.45 | 0.035 | -0.052 |
NEW | 0.262 | -0.289 | 0.489 | -0.405 | 0.523 | -0.252 | 0.322 | 0.058 | -0.013 |
NWN | 0.691 | -0.038 | 0.432 | -0.178 | -0.254 | 0.289 | -0.02 | -0.345 | 0.187 |
NWA | 0.715 | 0.04 | 0.38 | 0.071 | -0.383 | 0.031 | -0.07 | 0.408 | -0.14 |
CMDS | 0.722 | -0.317 | -0.351 | 0.226 | 0.151 | -0.033 | -0 | -0.215 | -0.365 |
CMDI | 0.637 | -0.343 | -0.434 | 0.248 | 0.267 | -0.014 | -0.07 | 0.161 | 0.354 |
EV | 2.269 | 1.494 | 1.295 | 1.039 | 0.956 | 0.661 | 0.579 | 0.383 | 0.323 |
PV | 25.21 | 16.60 | 14.39 | 11.55 | 10.63 | 7.34 | 6.44 | 4.25 | 3.59 |
CV | 25.21 | 41.81 | 56.20 | 67.75 | 78.37 | 85.72 | 92.15 | 96.41 | 100 |
Variable | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | PC7 | PC8 | PC9 |
|---|---|---|---|---|---|---|---|---|---|
PHT | -0.0810 | 0.3320 | 0.3990 | 0.5990 | 0.4640 | 0.3780 | 0.0760 | 0.0220 | -0.0290 |
LW | 0.3170 | 0.6530 | 0.1310 | 0.3300 | -0.0600 | -0.5650 | 0.0160 | -0.1310 | 0.0800 |
LBB | 0.3070 | 0.5720 | -0.4750 | -0.2130 | -0.0730 | 0.2070 | 0.5070 | 0.0570 | 0.0010 |
SP | 0.2890 | 0.5710 | -0.1120 | -0.4610 | 0.3910 | 0.0820 | -0.4500 | 0.0350 | -0.0520 |
NEW | 0.2620 | -0.2890 | 0.4890 | -0.4050 | 0.5230 | -0.2520 | 0.3220 | 0.0580 | -0.0130 |
NWN | 0.6910 | -0.0380 | 0.4320 | -0.1780 | -0.2540 | 0.2890 | -0.0190 | -0.3450 | 0.1870 |
NWA | 0.7150 | 0.0400 | 0.3800 | 0.0710 | -0.3830 | 0.0310 | -0.0660 | 0.4080 | -0.1400 |
CMDS | 0.7220 | -0.3170 | -0.3510 | 0.2260 | 0.1510 | -0.0330 | -0.0040 | -0.2150 | -0.3650 |
CMDI | 0.6370 | -0.3430 | -0.4340 | 0.2480 | 0.2670 | -0.0140 | -0.0730 | 0.1610 | 0.3540 |
EV | 2.27 | 1.49 | 1.30 | 1.04 | 0.96 | 0.66 | 0.58 | 0.38 | 0.32 |
PV | 25.21 | 16.60 | 14.39 | 11.55 | 10.63 | 7.34 | 6.44 | 4.25 | 3.59 |
CV | 25.21 | 41.81 | 56.20 | 67.75 | 78.37 | 85.72 | 92.15 | 96.41 | 100.00 |
Items | Degree of freedom (MAP) | Sum Sq (MAP) | Mean Sq (MAP) | F value (MAP) | Pr(>F) (MAP) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
MAPs | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 |
Lacewing | ||||||||||||||||||||
Block (ignoring Treatments) | 3 | 3 | 3 | 3 | 18.026 | 2.478 | 28.757 | 48.884 | 6.009 | 0.826 | 9.586 | 16.295 | 9.931 | 8.403 | 42.224 | 10.863 | 0.046 | 0.034 | 0.002 | 0.022 |
Treatment (eliminating Blocks) | 261 | 260 | 260 | 260 | 87.061 | 132.944 | 596.933 | 187.339 | 0.334 | 0.511 | 2.296 | 0.721 | 0.551 | 5.201 | 10.113 | 0.48 | 0.855 | 0.058 | 0.017 | 0.916 |
Treatment: Check | 1 | 1 | 1 | 1 | 0.005 | 1.176 | 0.68 | 5.12 | 0.005 | 1.176 | 0.68 | 5.12 | 0.008 | 11.96 | 2.997 | 3.413 | 0.933 | 0.026 | 0.158 | 0.138 |
Treatment: Test and Test vs. Check | 260 | 259 | 259 | 259 | 87.056 | 131.768 | 596.252 | 182.219 | 0.335 | 0.509 | 2.302 | 0.704 | 0.553 | 5.175 | 10.141 | 0.469 | 0.854 | 0.058 | 0.017 | 0.923 |
Residuals | 3 | 4 | 4 | 4 | 1.815 | 0.393 | 0.908 | 6 | 0.605 | 0.098 | 0.227 | 1.5 | ||||||||
Ladybird beetles | ||||||||||||||||||||
Block (ignoring Treatments) | 3 | 3 | 3 | 3 | 0.123 | 0.33 | 1.288 | 1.047 | 0.041 | 0.11 | 0.429 | 0.349 | 2.244 | 9.086 | 28.624 | 7.156 | 0.262 | 0.029 | 0.004 | 0.044 |
Treatment (eliminating Blocks) | 261 | 260 | 260 | 260 | 5.512 | 16.715 | 8.572 | 14.25 | 0.021 | 0.064 | 0.033 | 0.055 | 1.152 | 5.315 | 2.198 | 1.124 | 0.542 | 0.056 | 0.231 | 0.529 |
Treatment: Check | 1 | 1 | 1 | 1 | 0.405 | 0.027 | 0.08 | 0.125 | 0.405 | 0.027 | 0.08 | 0.125 | 22.091 | 2.254 | 5.333 | 2.564 | 0.018 | 0.208 | 0.082 | 0.185 |
Treatment: Test and Test vs. Check | 260 | 259 | 259 | 259 | 5.107 | 16.688 | 8.492 | 14.125 | 0.02 | 0.064 | 0.033 | 0.055 | 1.071 | 5.327 | 2.186 | 1.119 | 0.575 | 0.055 | 0.233 | 0.532 |
Residuals | 3 | 4 | 4 | 4 | 0.055 | 0.048 | 0.06 | 0.195 | 0.018 | 0.012 | 0.015 | 0.049 | ||||||||
Items | Degree of freedom (MAP) | Sum Sq (MAP) | Mean Sq (MAP) | F value (MAP) | Pr(>F) (MAP) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
MAPs | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 | 3 | 6 | 9 | 12 |
Block (ignoring Treatments | 3 | 3 | 3 | 3 | 1.114 | 0.23 | 4.67 | 6.51 | 0.371 | 0.077 | 1.557 | 2.17 | 18.575 | 1.387 | 11.974 | 64.295 | 0.019 | 0.368 | 0.018 | 0.001 |
Treatment (eliminating Blocks) | 261 | 260 | 260 | 260 | 35.566 | 30.509 | 48.812 | 42.302 | 0.136 | 0.117 | 0.188 | 0.163 | 6.813 | 2.118 | 1.444 | 4.821 | 0.068 | 0.244 | 0.402 | 0.066 |
Treatment: Check | 1 | 1 | 1 | 1 | 0.02 | 0.201 | 0.02 | 0.405 | 0.02 | 0.201 | 0.02 | 0.405 | 1 | 3.622 | 0.154 | 12 | 0.391 | 0.13 | 0.715 | 0.026 |
Treatment: Test and Test vs. Check | 260 | 259 | 259 | 259 | 35.546 | 30.308 | 48.792 | 41.897 | 0.137 | 0.117 | 0.188 | 0.162 | 6.836 | 2.112 | 1.449 | 4.793 | 0.068 | 0.245 | 0.401 | 0.066 |
Residuals | 3 | 4 | 4 | 4 | 0.06 | 0.222 | 0.52 | 0.135 | 0.02 | 0.055 | 0.13 | 0.034 | ||||||||
CMD | Cassava Mosaic Disease |
IPM | Integrated Pest Management |
MAP | Month After Planting |
PCA | Principal Component Analysis |
SLICASS | Sierra Leone Improved Cassava |
| [1] | Legg, J., Owor, B., Sseruwagi, P., and Ndunguru, J. Cassava mosaic virus disease in East and Central Africa: epidemiology and management of a regional pandemic. Advances in Virus Research, 2006, 67, pp. 355–418. |
| [2] | Patil, B., Legg, J., Kanju, E. and Fauquet, C. Cassava brown streak disease: a threat to food security in Africa. Journal of General Virology, 2015, 96, 956–968. |
| [3] | Nelson, S. Sooty Mold. Plant Disease PD-52. Mänoa, Honolulu, Hawai‘i, Cooperative extension Service, College of Tropical Agriculture and Human Resources, University of Hawaii, 2008. |
| [4] | Maruthi, M. N., Colvin, J., Seal, S. and Thresh, J. M. First report of a distinct begomovirus infecting cassava from Zanzibar. Plant Disease, 2002a, 86, 187. |
| [5] | Maruthi, M. N., Colvin, J., Seal, S., Gibson, G. and Cooper, J. Co-adaptation between cassava mosaic gemini viruses and their local vector populations. Virus Research, 86, 2002b, 71–85. |
| [6] | Thresh, J. M., Otim-Nape, G. W., Legg, J. P. and Fargette, D. African cassava mosaic virus disease: the magnitude of the problem. African Journal of Root and Tuber Crops, 1997, 2, 13. |
| [7] | Albajes, R., Sarasúa, M. J., Avilla, J., Arnó, J. and Gabarra, R. Integrated pest management in a Mediterranean region: the case of Catalonia, Spain. In Integrated Pest Management in the Global Arena, ed. KM Maredia, D Dakouo, D Mota-Sánchez, Oxon, UK: CABI Publishing, 2003, pp. 341–355. |
| [8] | Naranjo, S. E. and Ellsworth, P. C. Mortality dynamics and population regulation in Bemisia tabaci. Entomologia Experimentalis et Applicata, 2005, 116 (2), 93–108. |
| [9] | Gould, J., Hoelmer, K. A., and Goolsby, J. A. (eds). Classical Biological Control of Bemisia tabaci in the United States– A Review of Interagency Research and Implementation, Vol. 4, 344pp. Amsterdam, The Netherlands: Springer, 2008. |
| [10] | Obeng-Ofori, D., Owusu, E. O., and Afreh-Nuamah, K. Functional responses of lacewing predators to whitefly nymph densities. Journal of Biological Control, 1998, 12, 85–92. |
| [11] | Onzo, A., Hanna, R., and Zannou, I. Role of generalist predators in suppressing whitefly populations in cassava agroecosystems. Biological Control, 2009, 51(2), 123–131. |
| [12] | Hodek, I., and Honěk, A. Scale of polyphagy in ladybird beetles: Implications for biological control. Biological Control, 2009, 51(3), 439–447. |
| [13] | Nomikou, M., Janssen, A., Schraag, R., Sabelis, M. W. Phytoseiid predators as potential biological control agents for Bemisia tabaci. Experimental and Applied Acarology, 2001, 25, 271–291. |
| [14] | Naranjo, S. E., Ellsworth, P. C., Hagler, J. R. Conservation of natural enemies in cot ton: role of insect growth regulators for management of Bemisia tabaci. Biological Control, 2004b, 30, 52–72. |
| [15] | Naranjo, S. E., Cañas, L., and Ellsworth, P. C. Mortality and population dynamics of Bemisia tabaci within a multi-crop system. In Proceedings of the Third International Symposium on Biological Control of Arthropods, Christchurch, New Zealand, ed. PG Mason, DR Gillespie, CD Vincent, 2009, pp. 202–207. Publ. FHTET-2008-06: USDA Forest Service. |
| [16] | Macfadyen, S., Paull, C., Boykin, L. M., De Barro, P., Maruthi, M. N., Otim, M., Kalyebi, A., Vassão, D. G., Sseruwagi, P., Tay, W. T., Delatte, H., Seguni, Z., Colvin, J. and Omongo, C. A. Cassava whitefly, Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) in East African farming landscapes: a review of the factors determining abundance. Bulletin of Entomological Research, 2018, pp. 1–18. |
| [17] | Legg, J. P., Sseruwagi, P., Boniface, S., Okao-Okuja, G., Shirima, R., Bigirimana, S., Gashaka, G., Herrmann, H. W., Jeremiah, S., Obiero, H. Spatio-temporal patterns of genetic change amongst populations of cassava Bemisia tabaci whiteflies driving virus pandemics in East and Central Africa. Virus Research, 2014, 186, 61–75. |
| [18] | Akinlosotu, T. A., Jackai, L. E. N., and Adebayo, A. A. Whitefly infestations and cassava morphology: Implications for vector colonization dynamics. Journal of Tropical Agriculture, 1993, 70, 45–52. |
| [19] | Legg, J. P., and Zerihun, A. Seasonal Dynamics of Cassava Mosaic Disease Spread in East African Agroecosystems. Plant Disease, 2023, 107, 1451–1460. |
| [20] | Mbewe, N., Chipabika, G. and Mwila, M. Microenvironmental Drivers of Whitefly and Natural Enemy Dynamics in Cassava-Based Landscapes. Agriculture, Ecosystem and Environment, 2021a, 319, 107559. |
| [21] | Yila, K. M., Gboku, M. L. S., Lebbie, M. S. and Kamara, L. I. Changes in Rainfall and Temperature and Its Impact on Crop Production in Moyamba District, Southern Sierra Leone. Atmospheric and Climate Sciences, 2023, 13, 19-43. |
| [22] | Singh, K. and Varma, A. Modern Statistical Modeling Approaches in Vector Ecology and Entomology. Journal of Insect Science, 2023, 23, 1–15. |
| [23] | Abdullahi, A., Sani, I., Ibrahim, N. D. and Yusuf, I. Field Sampling Methods for Whitefly Population Assessment in Tropical Cropping Systems. Journal of Applied Entomology, 2021, 145, 795–804. |
| [24] | Diehl, J. W., Ellsworth, P. C. and Naranjo, S. E. Evaluation of a leaf turnmet hod for sampling white flies in cotton. In: Cotton, a College of Agriculture Report. The University of Arizona Cooperative Extension, Series P-99. 1995, pp. 241-246. |
| [25] | Kumar, P., Singh, R. and Arya, M. Field identification keys for predatory arthropods in tropical agricultural systems. Entomologia Experimentalis et Applicata, 2022, 170(3), 215–229. |
| [26] | Sseruwagi, P., Sserubombwe, W. S., Legg, J. P., Ndunguru, J. and Thresh, J. M. Methods of Surveying the Incidence and Severity of Cassava Mosaic Disease and Whitefly Vector Populations on Cassava in Africa: A Review. Virus Research, 2004, 100(1), 129–142. |
| [27] | Mwangi, P., Cheruiyot, D., and Kiplagat, O. Plant height and architectural influences on whitefly colonization in cassava. Tropical Plant Pathology, 2020, 45(6), 654–662. |
| [28] | Resende, M. D. V. Biometric Genetics and Statistics in the Breeding of Perennial Plants; Embrapa: Brasília, DF, Brazil, 2002. |
| [29] |
Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses: R Package Version 1.0.5.999. 2017. Available online:
http://www.sthda.com/english/rpkgs/factoextra (accessed on 9 December 2020). |
| [30] | Ratner, B. The correlation coefficient: Its values range between +1/−1, or do they?” Journal of Targeting, Measurement and Analysis for Marketing, 2009, 17(2), 139–142. |
| [31] | Singh, K., and Varma, A. Modern statistical approaches for entomological research: A practical guide. Journal of Insect Science, 2023, 23(2), 1–15. |
| [32] | Otim-Nape, G. W., Arah, R. O., and Kamau, J. Cassava whitefly population trends and virus transmission dynamics in Eastern Africa. African Crop Science Journal, 2021, 29(1), 57–74. |
| [33] | Mohammed, R., Olalekan, O., and Katungi, E. Ecological interactions shaping cassava pest dynamics under climate variability in sub-Saharan Africa. Journal of Applied Entomology, 2022, 146(4), 456–468. |
| [34] | Horowitz, A. R., and Gerling, D. Whitefly–natural enemy interactions: Biological and behavioral perspectives. Phytoparasitica, 1992, 20(1), 47–55. |
| [35] | Aregbesola, O. A., Sife, A. S., and Sæthre, M. G. Cassava canopy structure influences population dynamics of whiteflies and natural enemies. International Journal of Pest Management, 2020, 66(4), 345–357. |
| [36] | Mengistu, A., Wegary, D., and Woldehawariat, Y. Cassava canopy and microclimate characteristics influencing Bemisia tabaci distribution. Crop Protection, 2022, 156, 105954. |
| [37] | Santos, D., Ribeiro, M., and Silva, J. Diet breadth and predation efficiency of coccinellid beetles in tropical agroecosystems. Insects, 2021, 12(9), 789. |
| [38] | Kansiime, M., Nakato, V., and Alicai, T. Longitudinal dynamics of cassava mosaic disease symptom progression under varying whitefly pressure. Virus Research, 2023, 323, 199059. |
| [39] | FAO. Integrated Pest Management Guidelines for Sustainable Agriculture. Food and Agriculture Organization of the United Nations, 2022. |
| [40] | Kuntzman, J. and Brom, J. (2025). From Fields to Microclimate: Assessing the Influence of Agricultural Landscape Structure on Vegetation Cover and Local Climate in Central Europe. Remote Sensing, 2025, 17(1), 6. |
APA Style
Johnson, R. A. B., Samura, A. E., Bah, M. A., Cruz, I., Obeng-Ofori, D., et al. (2026). Whitefly–Natural Enemy Dynamics and Cassava Mosaic Disease Evaluated Under Field Condition in Sierra Leone. American Journal of Entomology, 10(1), 1-15. https://doi.org/10.11648/j.aje.20261001.11
ACS Style
Johnson, R. A. B.; Samura, A. E.; Bah, M. A.; Cruz, I.; Obeng-Ofori, D., et al. Whitefly–Natural Enemy Dynamics and Cassava Mosaic Disease Evaluated Under Field Condition in Sierra Leone. Am. J. Entomol. 2026, 10(1), 1-15. doi: 10.11648/j.aje.20261001.11
AMA Style
Johnson RAB, Samura AE, Bah MA, Cruz I, Obeng-Ofori D, et al. Whitefly–Natural Enemy Dynamics and Cassava Mosaic Disease Evaluated Under Field Condition in Sierra Leone. Am J Entomol. 2026;10(1):1-15. doi: 10.11648/j.aje.20261001.11
@article{10.11648/j.aje.20261001.11,
author = {Raymonda Adeline Bernardette Johnson and Alusaine Edward Samura and Mohamed Allieu Bah and Ivan Cruz and Daniel Obeng-Ofori and Paul Musa Lahai and Prince Emmanuel Norman},
title = {Whitefly–Natural Enemy Dynamics and Cassava Mosaic Disease Evaluated Under Field Condition in Sierra Leone},
journal = {American Journal of Entomology},
volume = {10},
number = {1},
pages = {1-15},
doi = {10.11648/j.aje.20261001.11},
url = {https://doi.org/10.11648/j.aje.20261001.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.aje.20261001.11},
abstract = {Bemisia tabaci is a major pest of cassava in sub-Saharan Africa, causing yield losses through direct feeding and its role in transmitting cassava mosaic disease (CMD). Natural enemies such as lacewings, ladybird beetles, and spiders provide valuable biological control services, yet their interactions with different whitefly developmental stages and plant structural traits remain insufficiently characterized. This study examined the dynamics among natural enemies, whitefly eggs, nymphs, adults, and plant height across 3, 6, 9, and 12 months after planting (MAP) under field conditions. The trial was conducted under natural cassava production conditions during 2020/2021 cropping season at the upland experimental site of the School of Agriculture and Food Sciences, Njala University. A total of 270 cassava genotypes comprising 268 local varieties and 2 improved checks (SLICASS 4 and SLICASS 6) were laid out in an augmented randomized design with four blocks. Results showed that lacewings and spiders strongly tracked nymph and adult whitefly populations, while ladybird beetles showed weaker associations. Principal Component Analysis (PCA) revealed alignment of predators with pest pressure during mid- and late season, whereas plant height exhibited minimal influence. Findings underscore the central role of lacewings and spiders in early and sustained suppression of whitefly populations, highlighting the importance of conservation-based integrated pest management (IPM) strategies. Findings serve as useful guide for conservation biological control as a primary IPM strategy for the enhancement of habitats for effective predators (lacewings and spiders) of the whitefly through reduced pesticide use, ground vegetation retention, intercropping, and maintenance of natural refuge habitats.},
year = {2026}
}
TY - JOUR T1 - Whitefly–Natural Enemy Dynamics and Cassava Mosaic Disease Evaluated Under Field Condition in Sierra Leone AU - Raymonda Adeline Bernardette Johnson AU - Alusaine Edward Samura AU - Mohamed Allieu Bah AU - Ivan Cruz AU - Daniel Obeng-Ofori AU - Paul Musa Lahai AU - Prince Emmanuel Norman Y1 - 2026/01/19 PY - 2026 N1 - https://doi.org/10.11648/j.aje.20261001.11 DO - 10.11648/j.aje.20261001.11 T2 - American Journal of Entomology JF - American Journal of Entomology JO - American Journal of Entomology SP - 1 EP - 15 PB - Science Publishing Group SN - 2640-0537 UR - https://doi.org/10.11648/j.aje.20261001.11 AB - Bemisia tabaci is a major pest of cassava in sub-Saharan Africa, causing yield losses through direct feeding and its role in transmitting cassava mosaic disease (CMD). Natural enemies such as lacewings, ladybird beetles, and spiders provide valuable biological control services, yet their interactions with different whitefly developmental stages and plant structural traits remain insufficiently characterized. This study examined the dynamics among natural enemies, whitefly eggs, nymphs, adults, and plant height across 3, 6, 9, and 12 months after planting (MAP) under field conditions. The trial was conducted under natural cassava production conditions during 2020/2021 cropping season at the upland experimental site of the School of Agriculture and Food Sciences, Njala University. A total of 270 cassava genotypes comprising 268 local varieties and 2 improved checks (SLICASS 4 and SLICASS 6) were laid out in an augmented randomized design with four blocks. Results showed that lacewings and spiders strongly tracked nymph and adult whitefly populations, while ladybird beetles showed weaker associations. Principal Component Analysis (PCA) revealed alignment of predators with pest pressure during mid- and late season, whereas plant height exhibited minimal influence. Findings underscore the central role of lacewings and spiders in early and sustained suppression of whitefly populations, highlighting the importance of conservation-based integrated pest management (IPM) strategies. Findings serve as useful guide for conservation biological control as a primary IPM strategy for the enhancement of habitats for effective predators (lacewings and spiders) of the whitefly through reduced pesticide use, ground vegetation retention, intercropping, and maintenance of natural refuge habitats. VL - 10 IS - 1 ER -
Department of Crop Protection, School of Agriculture and Food Sciences, Njala University, Njala Campus, Moyamba, Sierra Leone
Biography: Raymonda Adeline Bernardette Johnson PhD, Ministry of Agriculture and Food Security, Sierra Leone. Associate Lecturer, Crop Protection Department, Njala University. Expertise in biology, entomology, phytosanitary, pesticide management, IPM, integrated crop management, epidemiology, standards, food safety-CODEX, SPS, AMR, One health, trade and phytosanitary policy. She is a plant health technical experts, FAO and national consultant of WHO-CODEX program. An Africa representative in IPPC- Standard Committee. Currently the president of the West African Pesticide Registration Committee and deputy secretary of Toxicology and Ecotoxicology sub- committee. She was part of the committee that spear headed the formation of the African Phytosanitary Programme in collaboration with USDA, IPPC and AUIAPSC. Vice chair of National Food Safety Taskforce and Chair of the National Pesticide Management, National IPM and Fall Armyworm and National SPS Committees. Member of the Regional and National AMR Committee, national CODEX committee and a vice chair at the regional CODEX pesticide committee.

Department of Crop Protection, School of Agriculture and Food Sciences, Njala University, Njala Campus, Moyamba, Sierra Leone
Biography: Alusaine Edward Samura PhD is the acting head of the Crop Protection Department at the School of Agriculture and Food Science, Njala University and the Country Director of the Central and West African Virus Epidemiology WAVE Sierra Leone. culminating after 20 years’ research. He is currently serving as a as secretary of the Bio efficacy sub- committee in the West African Pesticide Registration Committee serving. He is a master trainer of the African Phytosanitary Programme on advanced plant health science and state-of-the-art digital tools to better equip field staff and administrators. He was instrumental in incorporating plant health into the One Health Governance Manual of the Republic of Sierra Leone. He is a member of the variety release and registration committee, National Pesticide Management Committee, national taskforces on Fall Armyworm and cassava viruses, National Food Safety taskforce as well as the Research and Publication Committee, School of Agriculture, Njala University.

Department of Crop Science, School of Agriculture and Food Sciences, Njala University, Njala Campus, Moyamba, Sierra Leone
Biography: Mohamed Allieu Bah, Ph.D. in Crop Science, Deputy Vice Chancellor, Njala Campus. Research portfolio demonstrates a strong commitment to sustainable agriculture and crop improvement in Sierra Leone and West Africa. His work on heavy metal stress tolerance in plants has contributed to understanding plant adaptation mechanisms, while his extensive work on rice breeding and evaluation directly supports food security in the region. His development of numerous training manuals reflects his dedication to knowledge transfer and capacity building in agricultural communities.

Department of Entomology, Brazilian Agricultural Research Corporation, Sete Lagoas, Brazil
Biography: Ivan Cruz, Agronomist, Master and PhD in Entomology; MBA in Project Management; Scientific Researcher; Biological Management of Insect Pests - Embrapa (Brazilian Agricultural Research Corporation). Graduated in agronomic engineering (1973) and started to work (1974) in insect research by the Brazilian Agricultural Research Corporation (EMBRAPA). Master's degree (1980) from Purdue University, USA, and a PhD from Brazilian University USP (1986) and Specialist degree in Project Management (MBA, 2021). He was president of the Brazilian Maize and Sorghum Association (ABMS) and is currently Editor of the International Journal of Maize and Sorghum. In terms of management, he was a General Manager at Embrapa Maize and Sorghum. At the national level of Embrapa, he was president of the Bio Inputs Research Portfolio Program. He participated in international research programs representing Embrapa. Recently (2022) he published the book “biological control of corn pests: an opportunity for farmers (Portuguese and English).

Department of Conservation Biology, Catholic University of Ghana, Sunyani, Ghana
Biography: Daniel Obeng-Ofori is a Professor of Applied Biology and Entomology and a distinguished University Administrator. He holds Bachelors in Agriculture, MPhil in Applied Biology and PhD in Entomology. Daniel won the prestigious Alexander von Humboldt Research Fellowship in Germany. He joined University of Ghana and became Professor in 2004. He has held several academic positions at Legon, UENR and CUG including Pro Vice-Chancellor and Vice-Chancellor. Daniel has conducted research and published widely and mentored several Bachelors, Masters and PhD scholars. He reviews scientific manuscripts for publication in 53 scholarly journals. He had been a Team Leader in many funded international research projects. Daniel was among the top World Scientists’ 2021 and 2022 rankings. He is a recipient of ICIPE’s 50th Anniversary Achievement Award in Nairobi, Kenya, 2020 and Ghana’s Most Respected CEO 2023 (Private University). And was Humboldt Ambassador Scientist for Ghana from 2016-2019.

Department of Forestry and Tree Crops Improvement Program, Kenema Forestry and Tree Crops Research Centre, Sierra Leone Agricultural Research Institute, Kenema, Sierra Leone
Biography: Paul Musa Lahai is a PhD holder with specialty in Plant Breeding and Genetics at the Njala University, Crop Science Department. He completed his PhD in Crop Science (Plant Breeding and Genetics) from Njala University in October, 2025, and has Master of Philosophy in Entomology from the University of Ghana, Legon in 2012. Furthermore, he obtained another Master of Advanced Studies in Integrated Crop Management at the University of Neuchatel, Switzerland. He has participated in multiple international research collaboration projects in recent years. He currently serves as the head of Tree Crops of the Sierra Leone Agricultural Research Institute since November, 2019.

Department of Germplasm Enhancement and Seeds Systems, Sierra Leone Agricultural Research Institute, Freetown, Sierra Leone
Biography: Prince Emmanuel Norman (PhD) is a Chief Research Officer and Deputy Director General, Research, Technology and Innovation Development at Sierra Leone Agricultural Research Institute (SLARI). He completed his PhD in Plant Breeding from University of Ghana in 2019, and his Master of Science in Plant Breeding from the University of KwaZulu-Natal, South Africa in 2011. Recognized for his exceptional contributions, Rev. Dr. Prince Emmanuel Norman has been honored with the INV Awards 2025 International Research and Invention Award, and Academic Awards ACH Awards 2025, etc. Dr. Norman's research has primarily centered around the genetic improvement of staple crops, particularly the white Guinea yam, cassava, sweet potato, cocoyam, cereals, etc. as well as agronomy, tissue culture, physiology, pathology, crop protection, etc. He has participated in multiple international research collaboration projects in recent years. He currently serves as Reviewer of numerous publications, examiner and mentor to many young upcoming researchers.

Information