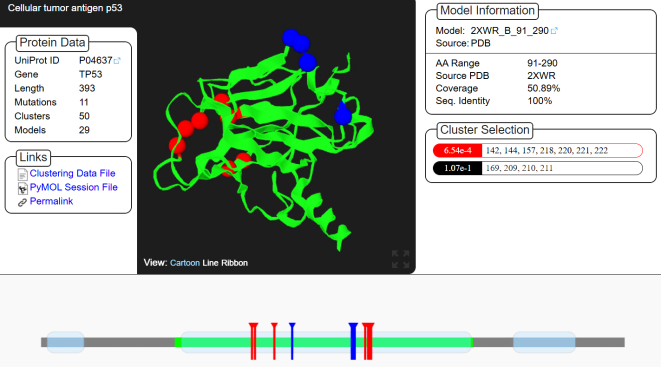
Prostate tumors are more common worldwide, with 60% of men over the age of 50 affected by benign prostatic hyperplasia (BPH) and 1.5 million new cases and 397,000 deaths from prostate cancer (PCa), which ranks as the second most common cancer globally. Although age is the most significant factor, other factors are associated with their development, and genetic factors appear to play a major role. This study aimed to evaluate the involvement of TP53 gene mutations in cases of prostate tumors among Senegalese men while contributing to the understanding of the mutational link between the two tumors. sixteen BPH tissue samples and seventeen PCa tissue samples were collected via biopsy from Senegalese patients following informed consent. DNA extraction followed by PCR amplification and sequencing were performed. Mutation Surveyor was used to identify mutations. Mutation Taster, Polyphen-2, SIFT, and SNP & GO were used to assess pathogenicity predictions. I-Mutant2, MuPro, and Dynamut2 were used to predict the stability, flexibility, and dynamics of the mutated p53 protein. MutPred2 and Mutation3D were used to predict physicochemical properties and map risk mutations. Variability, diversity, and genetic structure were determined using MEGA, BioEdit, DnaSP, and Arlequin. A total of 32 TP53 mutations were identified in the two tumors. These mutations were predominant in prostate cancer. No mutations shared between the two tumor types were found; however, shared mutations within each tumor type were observed, particularly one mutation (c.652G>A p.218Val>Met) present in all prostate cancer patients. Most non-synonymous mutations are predicted to be pathogenic and destabilizing for the mutated p53 protein in both tumors. Low polymorphism and a short genetic distance were observed between the two prostate tumors. This study provided insight into the potential impact of TP53 gene mutations on prostate tumors. Despite their low frequency in the Senegalese population, which may be explained by the small sample size, they play a role in the development and aggressiveness of prostate tumors. Therefore, special attention is required in patients carrying these mutations, particularly for the c.652G>A p.218Val>Met mutation, as the latter could influence management.
| Published in | International Journal of Genetics and Genomics (Volume 14, Issue 2) |
| DOI | 10.11648/j.ijgg.20261402.13 |
| Page(s) | 60-75 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Benign Prostatic Hyperplasia, Prostate Cancer, Mutations, TP53, Gene, Senegal
Tumour type | TP53 gene mutations | Number of patients with mutations | Total number (n) | Frequency of mutations (%) | |
|---|---|---|---|---|---|
Exon 5_Intron5 | Exon 6_Intron6 | ||||
Benign tumours | 1 | 10 | 4 | 16 | 25 |
Malignant tumours | 11 | 10 | 14 | 17 | 82.35 |
Mutations | Position | dbSNP | COSMIC | ClinVar | Effects of coding | Amino acid affected | Histology |
|---|---|---|---|---|---|---|---|
c.408A>G | Exon 5 | rs758781593 | COSM44154 | 185409 | Synonym | p.136Gln>Gln | Benign tumour |
c.424C>G | Exon 5 | New | COSM44969 | 3809688 | Non-synonymous | p.142Pro>Ala | Malignant tumour |
c.426T>A | Exon 5 | New | COSM44919 | New | Non-Synonym | p.142Pro>Ala | Malignant tumour |
c.431A>G | Exon 5 | New | COSM44028 | 2080519 | Non-synonymous | p.144Gln>Arg | Malignant tumour |
c.438G>A | Exon 5 | rs1131691026 | COSM10727 | 428890 | Nonsense | p.146Trp>* | Benign tumour |
c.469G>A | Exon 5 | rs121912654 | COSM43625 | 185404 | Non-synonymous | p.157Val>Ile | Malignant tumour |
c.506T>C | Exon 5 | New | COSM43851 | New | Non-synonymous | p.169Met>Thr | Malignant tumour |
c.559+13G>C | Intron 5 | New | New | New | No | Malignant tumour | |
c.559+27C>G | Intron 5 | rs778145407 | New | New | No | Malignant tumour | |
c.559+30G>A | Intron 5 | New | New | New | No | Malignant tumour | |
c.559+31G>C | Intron 5 | New | New | New | No | Malignant tumour | |
c.560-3T>G | Intron 5 | rs763746485 | COSM46059 | 634683 | No | Malignant tumour | |
c.626G>A | Exon 6 | New | COSM45995 | 962629 | Non-synonymous | p.209Arg>Lys | Benign tumour |
c.629A>G | Exon 6 | New | COSM45441 | 3232058 | Non-synonymous | p.210Asn>Ser | Benign tumour |
c.631A>G | Exon 6 | rs1060501198 | COSM44238 | 406581 | Non-synonymous | p.211Thr>Ala | Malignant tumour |
c.652G>A | Exon 6 | rs878854072 | COSM44683 | 237952 | Non-synonymous | p.218Val>Met | Malignant tumour |
c.654G>T | Exon 6 | New | New | 1754155 | Synonym | p.218Val>Val | Malignant tumour |
c.657C>G | Exon 6 | New | COSM44799 | New | Synonym | p.219Pro>Pro | Benign tumour |
c.659A>G | Exon 6 | rs121912666 | COSM10758 | 127819 | Non-synonymous | p.220Tyr>Cys | Benign tumour |
c.660T>G | Exon 6 | New | COSM44505 | New | Nonsense | p.220Y>* | Malignant tumour |
c.661G>T | Exon 6 | rs786201592 | COSM44817 | 634754 | Nonsense | p.221Glu>* | Malignant tumour |
c.663G>T | Exon 6 | New | COSM46369 | New | Non-synonymous | p.221Glu>Asp | Benign tumour |
c.665C>G | Exon 6 | New | New | New | Non-synonymous | p.222Pro>Arg | Malignant tumour |
c.666G>C | Exon 6 | New | COSM43924 | New | Synonym | p.222Pro>Pro | Benign tumour |
c.672+3C>A | Intron 6 | New | COSM6474332 | New | No | Malignant tumour | |
c.672+5G>C | Intron 6 | New | New | 2102775 | No | Malignant tumour | |
c.672+8T>A | Intron 6 | New | New | New | No | Malignant tumour | |
c.672+13A>G | Intron 6 | New | New | 1354615 | No | Malignant tumour | |
c.672+29G>A | Intron 6 | New | New | New | No | Benign tumour | |
c.672+30G>A | Intron 6 | rs200372146 | New | New | No | Benign tumour | |
c.672+31A>G | Intron 6 | rs34949160 | COSM45453 | 133420 | No | Benign tumour | |
c.672+32G>A | Intron 6 | New | New | New | No | Benign tumour |
Amino acids affected | Mutation Taster (score) | Polyphen-2 (score) | SIFT (score) | SNPs & GO | Histology |
|---|---|---|---|---|---|
c.424C>G p.142Pro>Ala | Deleterious (27) | Probably damaging (1) | Deleterious (0.02) | Diseased (0.820) | Malignant tumour |
c.431A>G p.144Gln>Arg | Deleterious (43) | Probably damaging (0.996) | Tolerated (0.08) | Neutral (0.307) | Malignant tumour |
c.469G>A p.157Val>Ile | Deleterious (29) | Potentially damaging (0.690) | Tolerated (0.54) | Neutral (0.043) | Malignant tumour |
c.506T>C p.169Met>Thr | Deleterious (81) | Benign (0.029) | Deleterious (0.02) | Diseased (0.623) | Malignant tumour |
c.626G>A p.209Arg>Lys | Deleterious (26) | Benign (0.00) | Tolerated (0.94) | Neutral (0.377) | Benign tumour |
c.629A>G p.210Asn>Ser | Benign (46) | Benign (0.276) | Tolerated (0.41) | Neutral (0.427) | Benign tumour |
c.631A>G p.211Thr>Ala | Deleterious (58) | Probably damaging (0.979) | Deleterious (0.01) | Diseased (0.782) | Malignant tumour |
c.652G>A p.218Val>Met | Deleterious (21) | Probably damaging (0.999) | Deleterious (0.00) | Diseased (0.822) | Malignant tumour |
c.659A>G p.220Tyr>Cys | Deleterious (194) | Potentially damaging (0.701) | Deleterious (0.00) | Diseased (0.886) | Benign tumour |
c.663G>T p.221Glu>Asp | Deleterious (45) | Benign (0.138) | Deleterious (0.00) | Diseased (0.833) | Benign tumour |
c.665C>G p.222Pro>Arg | Deleterious (103) | Potentially damaging (0.701) | Tolerated (0.09) | Neutral (0.489) | Malignant tumour |
Mutations | % BT | % MT |
|---|---|---|
c.424C>G p.142Pro>Ala | 0 | 9 |
c.431A>G p.144Gln>Arg | 0 | 9 |
c.469G>A p.157Val>Ile | 0 | 9 |
c.506T>C p.169Met>Thr | 0 | 45.45 |
c.626G>A p.209Arg>Lys | 9.09 | 0 |
c.629A>G p.210Asn>Ser | 9.09 | 0 |
c.631A>G p.211Thr>Ala | 0 | 27.27 |
c.652G>A p.218Val>Met | 0 | 100 |
c.659A>G p.220Tyr>Cys | 9.09 | 0 |
c.663G>T p.221Glu>Asp | 9.09 | 0 |
c.665C>G p.222Pro>Arg | 0 | 63.63 |
Variants | I-Mutant2.0 (PDB, 1TUP, Chain A) | MUpro | ||||
|---|---|---|---|---|---|---|
Stability | ΔΔG (kcal/mol) | Stability | ΔΔG (kcal/mol) | Stability | Confidence score | |
P142A | Decreased | -1.94 | Decreased | -0.371 | Decreased | -1 |
Q144R | Decreased | -1.22 | Decreased | -0.813 | Decreased | -0.199 |
V157I | Decreased | -1.50 | Decreased | -0.377 | Decreased | -0.283 |
M169T | Decreased | -0.72 | Decreased | -1.215 | Increased | 0.282 |
R209K | Decreased | -0.73 | Decreased | -1.524 | Decreased | -1 |
N210S | Decreased | -1.37 | Decreased | -1.243 | Decreased | -0.696 |
T211A | Decreased | -0.71 | Decreased | -1.299 | Decreased | -1 |
V218M | Decreased | -2.78 | Decreased | -0.707 | Decreased | -0.397 |
Y220C | Decreased | -1.60 | Decreased | -0.244 | Decreased | -0.771 |
E221N | Decreased | -0.15 | Decreased | -0.823 | Decreased | -1 |
P222R | Decreased | -1.19 | Decreased | -0.878 | Decreased | -0.427 |
Variants | ΔΔG (kcal/mol) | Protein stability | Wild Type | Mutant |
|---|---|---|---|---|
P142A | -2,06 | Destabilising |
|
|
Q144R | -0,32 | Destabilising |
|
|
V157I | -0,95 | Destabilising |
|
|
M169T | -0,43 | Destabilising |
|
|
R209K | -0,08 | Destabilising |
|
|
N210S | 0,14 | Stabilising |
|
|
T211A | -1,08 | Destabilising |
|
|
V218M | -0,92 | Destabilising |
|
|
Y220C | -2,37 | Destabilising |
|
|
E221N | -0,65 | Destabilising |
|
|
P222R | -0,04 | Destabilising |
|
|
Variants | MutPre2 score | Molecular mechanisms | P-value |
|---|---|---|---|
P142A | 0.607 | Altered transmembrane protein | 0.03 |
Gain of disulphide bond at C141 | 0.05 | ||
Q144R | 0.413 | - | - |
V157I | 0.162 | - | - |
M169T | 0.288 | - | - |
R209K | 0.067 | - | - |
N210S | 0.065 | - | - |
T211A | 0.446 | - | - |
V218M | 0.462 | - | - |
Y220C | 0.638 | Strand loss | 0.01 |
Altered transmembrane protein | 0.02 | ||
Loss of sulphation at Y220 | 0.05 | ||
E221N | 0.353 | - | - |
P222R | 0.207 | - | - |
Benign prostate tumours | Malignant prostate tumours | |||
|---|---|---|---|---|
Genetic variability parameters | ||||
Variables | Number | Percentage | Number | Percentage |
Number of individuals | 17 | 51.51% | 16 | 48.48% |
Number of sites | 350 | 350 | ||
Monomorphic sites | 345 | 98.57 | 338 | 96.57 |
Polymorphic sites | 5 | 1.42% | 12 | 3.43 |
Non-informative variable sites | 4 | 1.14% | 8 | 2.29 |
Variable sites in parsimony | 1 | 0.29 | 4 | 1.14 |
Total number of mutations (Eta) | 5 | 12 | ||
Average number of nucleotide differences (k) | 0.733 | 2.382 | ||
Percentage of transition | 35.7 | 58.14 | ||
Percentage of Transversion | 64.32 | 41.86 | ||
Mutation rate (R) | 0.519 | 1.298 | ||
Number of haplotypes | 4 | 11 | ||
Genetic diversity indices | ||||
Hd (Variance) | 0.350 ± 0.2183 | 0.882 ± 0.00516 | ||
Pi (variance) | 0.00210 ± 0.0000011 | 0.00681 ± 0.0000012 | ||
Amino acids | BT | MT | P-Value |
|---|---|---|---|
Ala | 11.494 | 11.596 | 8.092e-05*** |
Cys | 4.454 | 4.570 | 6.284e-05*** |
Asp | 1.149 | 1.160 | 1.066e-05*** |
Glu | 3.448 | 3.479 | 1.066e-05*** |
Phe | 1.149 | 1.228 | 3.452e-06*** |
Gly | 2.443 | 2.456 | 0.001813 |
His | 2.299 | 2.387 | 1.278e-05*** |
Ile | 6.897 | 7.026 | 1.278e-05*** |
Lys | 1.221 | 1.160 | 1.278e-05*** |
Leu | 9.195 | 9.277 | 1.066e-05*** |
Met | 4.598 | 4.570 | 0.000225*** |
Asn | 1.078 | 0.477 | 0.09862 |
Pro | 12.716 | 12.688 | 0.0009343*** |
Gln | 1.1494 | 1.160 | 1.066e-05*** |
Arg | 5.603 | 5.798 | 1.119e-05*** |
Ser | 12.643 | 13.302 | 0.0003191*** |
Thr | 5.819 | 5.799 | 0.0002292*** |
Val | 3.448 | 3.479 | 1.066e-05*** |
Trp | 9.195 | 8.390 | 1.066e-05*** |
Tyr | 0 | 0 | - |
Type of tissue | Intra-population distance ± standard deviation | Inter-population distance ± standard deviation | Fst (P-Value) | AMOVA test | |||
|---|---|---|---|---|---|---|---|
TB | TM | TB | TM | intra-population | inter-population | ||
TB | 0.00 ± 0.00 | 0.00391 ± 0.00265 | 0.37792 (0.00±0.000) | 62.21 | 37.79 | ||
TM | 0.01 ± 0.00 | ||||||
DNA | Deoxyribonucleic Acid |
dbSNP | Single Nucleotide Polymorphism Database |
dNTP | Deoxyribonucleotide Triphosphate |
COSMIC | Catalogue of Somatic Mutations in Cancer |
Fst | Fixation Index |
BPH | Benign Prostatic Hyperplasia |
NCBI | National Center for Biotechnology Information |
p53 | Protein 53 |
PCa | Prostate Cancer |
PCR | Polymerase Chain Reaction |
PDB | Protein Data Bank |
PSA | Prostate-Specific Antigen |
PSIC | Position-Specific Independent Counts |
SIFT | Sorting Intolerant from Tolerant |
SNP | Single Nucleotide Polymorphism |
TP53 | Tumor Protein p53 |
| [1] | Egan, K. B. (2016). The Epidemiology of Benign Prostatic Hyperplasia Associated with Lower Urinary Tract Symptoms : Prevalence and Incident Rates. Urologic Clinics of North America, 43(3), 289-297. |
| [2] | Gupta, S., Gupta, G., & L. Sharma, V. (2017). Evolving Novel Chemical Entities for Management of Benign Prostatic Hyperplasia#. Mini-Reviews in Medicinal Chemistry, 17(7), 593-602. |
| [3] | National Agency for Statistics and Demography (ANSD). Report on Population Projections for Senegal, 2023–2073. Accessed April 13, 2026, at |
| [4] | Bray, F., Laversanne, M., Sung, H., Ferlay, J., Siegel, R. L., Soerjomataram, I., & Jemal, A. (2024). Global cancer statistics 2022 : GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer Journal for Clinicians, 74(3), 229-263. |
| [5] | Milliron, B.-J., Bruneau, M., Obeid, E., Gross, L., Bealin, L., Smaltz, C., & Giri, V. N. (2019). Diet assessment among men undergoing genetic counseling and genetic testing for inherited prostate cancer : Exploring a teachable moment to support diet intervention. The Prostate, 79(7), 778-783. |
| [6] | Baker, S. J., & Vogelstein, B. (2019). p53 : A tumor suppressor hiding in plain sight. Journal of Molecular Cell Biology, 11(7), 536-538. |
| [7] | Olivier, M., Hollstein, M., & Hainaut, P. (2010). TP53 Mutations in Human Cancers : Origins, Consequences, and Clinical Use. Cold Spring Harbor Perspectives in Biology, 2(1), a001008. |
| [8] | Rozas, J., Ferrer-Mata, A., Sánchez-DelBarrio, J. C., Guirao-Rico, S., Librado, P., Ramos-Onsins, S. E., & Sánchez-Gracia, A. (2017). DnaSP 6 : DNA Sequence Polymorphism Analysis of Large Data Sets. Molecular Biology and Evolution, 34(12), 3299-3302. |
| [9] | Tamura, K., Stecher, G., & Kumar, S. (2021). MEGA11 : Molecular Evolutionary Genetics Analysis Version 11. Molecular Biology and Evolution, 38(7), 3022-3027. |
| [10] | Excoffier, L., Laval, G., & Schneider, S. (2007). Arlequin (version 3.0) : An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online, 1, 47-50. |
| [11] | Zole, E., Baumanis, E., Freimane, L., Dāle, R., Leiše, A., Lietuvietis, V., & Ranka, R. (2024). Changes in TP53 Gene, Telomere Length, and Mitochondrial DNA in Benign Prostatic Hyperplasia Patients. Biomedicines, 12(10), Article 10. |
| [12] | Schlechte, H., Lenk, S. V., Löning, T., Schnorr, D., Rudolph, B. D., Ditscherlein, G., & Loening, S. A. (1998). P53 tumour suppressor gene mutations in benign prostatic hyperplasia and prostate cancer. European Urology, 34(5), 433-440. |
| [13] | Zhou, J., Lai, Y., Peng, S., Tang, C., Chen, Y., Li, L., Huang, H., & Guo, Z. (2022). Comprehensive analysis of TP53 and SPOP mutations and their impact on survival in metastatic prostate cancer. Frontiers in Oncology, 12, 957404. |
| [14] | Aboulalaa, K., Laraqui, A., Tagajdid, R., Ennibi, K., & Ennaji, M. M. (2024). Germline mutations of TP53 gene can be a key factor in prostate cancer genesis. Exploration of Medicine, 641-655. |
| [15] | Wang, Z., Burigotto, M., Ghetti, S., Vaillant, F., Tan, T., Capaldo, B. D., Palmieri, M., Hirokawa, Y., Tai, L., Simpson, D. S., Chang, C., Huang, A. S., Lieschke, E., Diepstraten, S. T., Kaloni, D., Riffkin, C., Huang, D. C. S., Li Wai Suen, C. S. N., Garnham, A. L., … Strasser, A. (2024). Loss-of-Function but Not Gain-of-Function Properties of Mutant TP53 Are Critical for the Proliferation, Survival, and Metastasis of a Broad Range of Cancer Cells. Cancer Discovery, 14(2), 362-379. |
| [16] | Petitjean, A., Mathe, E., Kato, S., Ishioka, C., Tavtigian, S. V., Hainaut, P., & Olivier, M. (2007). Impact of mutant p53 functional properties on TP53 mutation patterns and tumor phenotype : Lessons from recent developments in the IARC TP53 database. Human Mutation, 28(6), 622-629. |
| [17] | Du, X., Zheng, J., Lu, X., & Zhang, Y. (2025). A Truncated Mutation of TP53 Promotes Chemoresistance in Tongue Squamous Cell Carcinoma. International Journal of Molecular Sciences, 26(5), 2353. |
| [18] | Cho, Y., Gorina, S., Jeffrey, P. D., & Pavletich, N. P. (1994). Crystal structure of a p53 tumor suppressor-DNA complex : Understanding tumorigenic mutations. Science (New York, N. Y.), 265(5170), 346-355. |
| [19] | Kato, S., Han, S.-Y., Liu, W., Otsuka, K., Shibata, H., Kanamaru, R., & Ishioka, C. (2003). Understanding the function–structure and function–mutation relationships of p53 tumor suppressor protein by high-resolution missense mutation analysis. Proceedings of the National Academy of Sciences of the United States of America, 100(14), 8424-8429. |
| [20] | Gully, C. P., Velazquez-Torres, G., Shin, J.-H., Fuentes-Mattei, E., Wang, E., Carlock, C., Chen, J., Rothenberg, D., Adams, H. P., Choi, H. H., Guma, S., Phan, L., Chou, P.-C., Su, C.-H., Zhang, F., Chen, J.-S., Yang, T.-Y., Yeung, S.-C. J., & Lee, M.-H. (2012). Aurora B kinase phosphorylates and instigates degradation of p53. Proceedings of the National Academy of Sciences, 109(24). |
| [21] | Shan, L., Yu, J., He, Z., Chen, S., Liu, M., Ding, H., Xu, L., Zhao, J., Yang, A., & Jiang, H. (2020). Defining relative mutational difficulty to understand cancer formation. Cell Discovery, 6(1), 48. |
| [22] | McNeal, J. E., Redwine, E. A., Freiha, F. S., & Stamey, T. A. (1988). Zonal Distribution of Prostatic Adenocarcinoma: Correlation with Histologic Pattern and Direction of Spread. The American Journal of Surgical Pathology, 12(12), 897. |
| [23] | Berner, A., Geitvik, G., Karlsen, F., Fosså, S. D., Nesland, J. M., & BØRresen, A.-L. (1995). TP53 mutations in prostatic cancer. Analysis of pre- and post-treatment archival formalin-fixed tumour tissue. The Journal of Pathology, 176(3), 299-308. |
| [24] | Yang, F., Petsalaki, E., Rolland, T., Hill, D. E., Vidal, M., & Roth, F. P. (2015). Protein domain-level landscape of cancer-type-specific somatic mutations. PLoS Computational Biology, 11(3), e1004147. |
| [25] | Nachman, M. W., & Crowell, S. L. (2000). Estimate of the mutation rate per nucleotide in humans. Genetics, 156(1), 297-304. |
| [26] | Alexandrov, L. B., Kim, J., Haradhvala, N. J., Huang, M. N., Tian Ng, A. W., Wu, Y., Boot, A., Covington, K. R., Gordenin, D. A., Bergstrom, E. N., Islam, S. M. A., Lopez-Bigas, N., Klimczak, L. J., McPherson, J. R., Morganella, S., Sabarinathan, R., Wheeler, D. A., Mustonen, V., Getz, G., … Stratton, M. R. (2020). The repertoire of mutational signatures in human cancer. Nature, 578(7793), 94-101. |
| [27] | McGranahan, N., Favero, F., de Bruin, E. C., Birkbak, N. J., Szallasi, Z., & Swanton, C. (2015). Clonal status of actionable driver events and the timing of mutational processes in cancer evolution. Science translational medicine, 7(283), 283ra54. |
| [28] | Turajlic, S., & Swanton, C. (2016). Metastasis as an evolutionary process. Science, 352(6282), 169-175. |
| [29] | Ben-David, U., & Amon, A. (2020). Context is everything : Aneuploidy in cancer. Nature Reviews. Genetics, 21(1), 44-62. |
| [30] | Hussain, A., Xie, L., Deng, G., & Kang, X. (2023). Common alterations in plasma free amino acid profiles and gut microbiota-derived tryptophan metabolites of five types of cancer patients. Amino Acids, 55(9), 1189-1200. |
| [31] | ynakiewicz, A., Stanislawska-Sachadyn, A., Sawicka-Zukowska, M., Galezowska, G., Ratajczyk, J., Owczarzak, A., Skuza, M., Wolska, L., & Stachowicz-Stencel, T. (2021). Plasma free amino acid profiling as metabolomic diagnostic and prognostic biomarker in paediatric cancer patients : A follow-up study. Amino Acids, 53(1), 133-138. |
| [32] | Marusyk, A., Almendro, V., & Polyak, K. (2012). Intra-tumour heterogeneity : A looking glass for cancer ? Nature Reviews Cancer, 12(5), 323-334. |
| [33] | Ørsted, D. D., & Bojesen, S. E. (2013). The link between benign prostatic hyperplasia and prostate cancer. Nature Reviews Urology, 10(1), 49-54. |
| [34] | Kim, S. S., Lee, S. C., Lim, B., Shin, S.-H., Kim, M. Y., Kim, S.-Y., Lim, H., Charton, C., Shin, D., Moon, H. W., Kim, J., Park, D., Park, W.-Y., & Lee, J. Y. (2023). DNA methylation biomarkers distinguishing early-stage prostate cancer from benign prostatic hyperplasia. Prostate International, 11(2), 113-121. |
APA Style
Ahamada, F., Ndong, A., Samb, M. D., Mbaye, F., Sembene, M. (2026). Frequency and Characterization of TP53 Tumor Suppressor Gene Mutations in the Development and Progression of Prostate Tumors in Senegal. International Journal of Genetics and Genomics, 14(2), 60-75. https://doi.org/10.11648/j.ijgg.20261402.13
ACS Style
Ahamada, F.; Ndong, A.; Samb, M. D.; Mbaye, F.; Sembene, M. Frequency and Characterization of TP53 Tumor Suppressor Gene Mutations in the Development and Progression of Prostate Tumors in Senegal. Int. J. Genet. Genomics 2026, 14(2), 60-75. doi: 10.11648/j.ijgg.20261402.13
@article{10.11648/j.ijgg.20261402.13,
author = {Fahimat Ahamada and Anna Ndong and Mame Diarra Samb and Fatimata Mbaye and Mbacke Sembene},
title = {Frequency and Characterization of TP53 Tumor Suppressor Gene Mutations in the Development and Progression of Prostate Tumors in Senegal},
journal = {International Journal of Genetics and Genomics},
volume = {14},
number = {2},
pages = {60-75},
doi = {10.11648/j.ijgg.20261402.13},
url = {https://doi.org/10.11648/j.ijgg.20261402.13},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijgg.20261402.13},
abstract = {Prostate tumors are more common worldwide, with 60% of men over the age of 50 affected by benign prostatic hyperplasia (BPH) and 1.5 million new cases and 397,000 deaths from prostate cancer (PCa), which ranks as the second most common cancer globally. Although age is the most significant factor, other factors are associated with their development, and genetic factors appear to play a major role. This study aimed to evaluate the involvement of TP53 gene mutations in cases of prostate tumors among Senegalese men while contributing to the understanding of the mutational link between the two tumors. sixteen BPH tissue samples and seventeen PCa tissue samples were collected via biopsy from Senegalese patients following informed consent. DNA extraction followed by PCR amplification and sequencing were performed. Mutation Surveyor was used to identify mutations. Mutation Taster, Polyphen-2, SIFT, and SNP & GO were used to assess pathogenicity predictions. I-Mutant2, MuPro, and Dynamut2 were used to predict the stability, flexibility, and dynamics of the mutated p53 protein. MutPred2 and Mutation3D were used to predict physicochemical properties and map risk mutations. Variability, diversity, and genetic structure were determined using MEGA, BioEdit, DnaSP, and Arlequin. A total of 32 TP53 mutations were identified in the two tumors. These mutations were predominant in prostate cancer. No mutations shared between the two tumor types were found; however, shared mutations within each tumor type were observed, particularly one mutation (c.652G>A p.218Val>Met) present in all prostate cancer patients. Most non-synonymous mutations are predicted to be pathogenic and destabilizing for the mutated p53 protein in both tumors. Low polymorphism and a short genetic distance were observed between the two prostate tumors. This study provided insight into the potential impact of TP53 gene mutations on prostate tumors. Despite their low frequency in the Senegalese population, which may be explained by the small sample size, they play a role in the development and aggressiveness of prostate tumors. Therefore, special attention is required in patients carrying these mutations, particularly for the c.652G>A p.218Val>Met mutation, as the latter could influence management.},
year = {2026}
}
TY - JOUR T1 - Frequency and Characterization of TP53 Tumor Suppressor Gene Mutations in the Development and Progression of Prostate Tumors in Senegal AU - Fahimat Ahamada AU - Anna Ndong AU - Mame Diarra Samb AU - Fatimata Mbaye AU - Mbacke Sembene Y1 - 2026/05/12 PY - 2026 N1 - https://doi.org/10.11648/j.ijgg.20261402.13 DO - 10.11648/j.ijgg.20261402.13 T2 - International Journal of Genetics and Genomics JF - International Journal of Genetics and Genomics JO - International Journal of Genetics and Genomics SP - 60 EP - 75 PB - Science Publishing Group SN - 2376-7359 UR - https://doi.org/10.11648/j.ijgg.20261402.13 AB - Prostate tumors are more common worldwide, with 60% of men over the age of 50 affected by benign prostatic hyperplasia (BPH) and 1.5 million new cases and 397,000 deaths from prostate cancer (PCa), which ranks as the second most common cancer globally. Although age is the most significant factor, other factors are associated with their development, and genetic factors appear to play a major role. This study aimed to evaluate the involvement of TP53 gene mutations in cases of prostate tumors among Senegalese men while contributing to the understanding of the mutational link between the two tumors. sixteen BPH tissue samples and seventeen PCa tissue samples were collected via biopsy from Senegalese patients following informed consent. DNA extraction followed by PCR amplification and sequencing were performed. Mutation Surveyor was used to identify mutations. Mutation Taster, Polyphen-2, SIFT, and SNP & GO were used to assess pathogenicity predictions. I-Mutant2, MuPro, and Dynamut2 were used to predict the stability, flexibility, and dynamics of the mutated p53 protein. MutPred2 and Mutation3D were used to predict physicochemical properties and map risk mutations. Variability, diversity, and genetic structure were determined using MEGA, BioEdit, DnaSP, and Arlequin. A total of 32 TP53 mutations were identified in the two tumors. These mutations were predominant in prostate cancer. No mutations shared between the two tumor types were found; however, shared mutations within each tumor type were observed, particularly one mutation (c.652G>A p.218Val>Met) present in all prostate cancer patients. Most non-synonymous mutations are predicted to be pathogenic and destabilizing for the mutated p53 protein in both tumors. Low polymorphism and a short genetic distance were observed between the two prostate tumors. This study provided insight into the potential impact of TP53 gene mutations on prostate tumors. Despite their low frequency in the Senegalese population, which may be explained by the small sample size, they play a role in the development and aggressiveness of prostate tumors. Therefore, special attention is required in patients carrying these mutations, particularly for the c.652G>A p.218Val>Met mutation, as the latter could influence management. VL - 14 IS - 2 ER -
Department of Animal Biology, Cheikh Anta Diop University, Dakar, Senegal
Department of Animal Biology, Cheikh Anta Diop University, Dakar, Senegal
Department of Animal Biology, Cheikh Anta Diop University, Dakar, Senegal
Department of Animal Biology, Cheikh Anta Diop University, Dakar, Senegal
Department of Animal Biology, Cheikh Anta Diop University, Dakar, Senegal
Information