Iron ions (Fe2+ and Fe3+) are essential trace elements for the human body, and are often added to various foods, but their effects on protein glycation remain unclear. This study evaluated the differential influences of Fe2+ and Fe3+ on the glycation reaction of β-lactoglobulin (β-Lg)-D-ribose system in terms of glycation degree, protein conformation and the distribution of modification sites. Free amino group contents and HPLC HCD MS/MS analyses indicated that both Fe3+ and Fe2+ could catalyze the glycation process and increase the glycated sites. The system contain Fe2+ exhibited higher glycation degree and more glycation sites (8), and lesser glycation sites were identified in system contain Fe3+ (5) and system without ferric ions (2). Additional sites (L1, K14, K135) were facilitated glycation by Fe2+, and most glycation sites showed higher degree of substitution per peptide (DSP) values when with Fe2+. In comparison with Fe2+, Fe3+ caused more pronounced alterations on both secondary and tertiary protein structure, promoted the β-Lg unfolding, and changed the protein structure to a more unordered form. In conclusion, Fe2+ at a specified concentration was a better choice to promote glycation reaction while maintain the protein structure. This study provide a theoretical basis for protein glycation modification with iron ions at different valence states participated.
| Published in | International Journal of Nutrition and Food Sciences (Volume 15, Issue 3) |
| DOI | 10.11648/j.ijnfs.20261503.11 |
| Page(s) | 93-103 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
β-lactoglobulin, Ferric Ions, Glycation, Mass Spectrometry, Protein Structure
NO. | Samples | Secondary structure | |||
|---|---|---|---|---|---|
α-Helix (%) | β-Sheet (%) | β-Turns (%) | Unordered (%) | ||
1 | Natural β-Lg | 20.4±0.33f | 39.1±5.24c | 16.6±1.70a | 24.3±0.53a |
2 | β-control | 18.9±0.90e,f | 28.7±1.37b | 24.5±1.80c | 27.0±0.71b |
3 | β-R-control | 19.6±0.45e,f | 29.7±0.62b | 23.4±0.61b,c | 27.4±1.21b |
4 | β-R-0.01-Fe2+ | 16.6±1.04c,d | 28.6±1.55b | 22.1±0.25b | 32.0±1.01c,d |
5 | β-R-0.02-Fe2+ | 18.0±1.61d,e | 27.5±0.81a,b | 22.2±0.35b | 32.2±0.11c |
6 | β-R-0.03-Fe2+ | 16.1±1.34b,c | 30.1±1.62b | 21.8±0.24b | 30.1±1.44c |
7 | β-R-0.01-Fe3+ | 17.0±0.75c,d | 28.7±0.87b | 21.9±0.11b | 31.3±0.78c,d |
8 | β-R-0.02-Fe3+ | 13.9±1.15a | 23.9±3.04a | 23.3±0.45b,c | 42.3±2.25f |
9 | β-R-0.03-Fe3+ | 14.5±0.45a,b | 25.7±3.50a,b | 24.3±0.25c | 36.6±0.21e |
No. | m/z | Delta ppm | Start | End | Sequence |
|---|---|---|---|---|---|
1 | 609.8421+2 | 0.564 | 1 | 11 | (-)LIVTQTMKGLD(I) |
2 | 451.7583+2 | -0.116 | 12 | 19 | (D)IQKVAGTW(Y) |
3 | 584.275+1 | 0.581 | 20 | 24 | (W)YSLAM(A) |
4 | 694.308+1 | 0.407 | 24 | 30 | (A)MAASDIS(L) |
5 | 445.302+1 | -0.138 | 29 | 32 | (D)ISLL(D) |
6 | 478.7616+2 | -0.049 | 33 | 41 | (L)DAQSAPLRV(Y) |
7 | 652.32+1 | 1.94 | 42 | 46 | (V)YVEEL(K) |
8 | 571.3081+1 | -0.881 | 47 | 51 | (L)KPTPE(G) |
9 | 885.4566+1 | 0.237 | 50 | 57 | (T)PEGDLEIL(L) |
10 | 502.2462+2 | 0.785 | 58 | 65 | (L)LQKWENGE(C) |
11 | 700.472+1 | 0.59 | 67 | 72 | (C)AQKKII(A) |
12 | 460.2766+1 | 0.0543 | 72 | 75 | (I)IAEK(T) |
13 | 452.2865+2 | -0.544 | 75 | 82 | (E)KTKIPAVF(K) |
14 | 401.7190+2 | 0.736 | 83 | 89 | (F)KIDALNE(N) |
15 | 685.4608+1 | 0.164 | 90 | 95 | (E)NKVLVL(D) |
16 | 579.8058+2 | 0.179 | 95 | 103 | (V)LDTDYKKYL(L) |
17 | 555.318+1 | -0.0208 | 102 | 105 | (K)YLLF(C) |
18 | 783.265+1 | 0.809 | 106 | 112 | (F)CMENSAE(P) |
19 | 573.288+1 | 0.0558 | 113 | 117 | (E)PEQSL(V) |
20 | 678.3308+1 | -0.781 | 117 | 122 | (S)LVCQCL(V) |
21 | 408.2165+2 | -0.0428 | 123 | 129 | (L) VRTPEVD (D) |
22 | 483.7243+2 | -0.15 | 130 | 137 | (D)DEALEKFD(K) |
23 | 460.7820+2 | 0.375 | 135 | 142 | (E)KFDKALKA(L) |
24 | 440.2655+1 | 0.477 | 143 | 149 | (A)LPMHIRL(S) |
25 | 806.4048+1 | 0.627 | 150 | 156 | (L)SFNPTQL(E) |
26 | 758.3143+1 | 0.705 | 157 | 162 | (L)EEQCHI(-) |
m/z | Delta ppm | Start | End | Sequence | Glycated site | Modified peptide | DSP |
|---|---|---|---|---|---|---|---|
β-R-0.03-Fe2+ | |||||||
609.8421+2 | 0.564 | 1 | 11 | (-)LIVTQTMKGLD(I) | L1, K8 | 675.8632+2/741.8890+2 | 4.5% |
451.7583+2 | -0.116 | 12 | 19 | (D)IQKVAGTW(Y) | K14 | 517.8100+2 | 2.5% |
571.3081+1 | -0.881 | 47 | 51 | (L)KPTPE(G) | K47 | 703.3492+1 | 56.01% |
502.2462+2 | 0.785 | 58 | 65 | (L)LQKWENGE(C) | K60 | 568.2684+2 | 42.46% |
401.7190+2 | 0.736 | 83 | 89 | (F)KIDALNE(N) | K83 | 467.7312+2 | 53.03% |
408.2165+2 | -0.0428 | 123 | 129 | (L) VRTPEVD (D) | R124 | 474.2560+2 | 41.38% |
607.3456+1 | 0.0543 | 132 | 135 | (E)ALEK(L) | K135 | 739.3869+1 | 5.51% |
β-R-0.03-Fe3+ | |||||||
609.8417+2 | -0.0921 | 1 | 11 | (-)LIVTQTMKGLD(I) | K8 | 678.8641+2 | 3.95% |
571.3081+1 | -0.881 | 47 | 51 | (L)KPTPE(G) | K47 | 703.3351+1 | 1.95% |
502.2460+2 | 0.386 | 58 | 65 | (L)LQKWENGE(C) | K60 | 568.2678+2 | 31.64% |
401.7190+2 | 0.267 | 83 | 89 | (F)KIDALNE(N) | K83 | 467.7298+2 | 41.75% |
408.2164+2 | -0.836 | 123 | 129 | (L)VRTPEVD(E) | R124 | 474.2557+2 | 61.71% |
β-R-control | |||||||
609.8418+2 | 0.072 | 1 | 11 | (-)LIVTQTMKGLD(I) | K8 | 675.8632+2 | 4.41% |
579.8057+2 | 0.00588 | 95 | 103 | (V)LDTDYKKYL(L) | K100 | 645.8271+2 | 1.43% |
β-Lg | β-Lactoglobulin |
LC-HRMS | Liquid Chromatography High-Resolution Mass Spectrometry |
UV | Ultraviolet Spectrometry |
CD | Circular Dichroism |
DTT | DL-Dithiothreitol |
OPA | o-Phthalaldehyde |
DSP | Degree of Substitution per Peptide |
HCD | High-energy C-trap Dissociation |
FA | Formic Acid |
| [1] | Cheng, Y. H., Tang, W. J., Xu, Z., Wen, L., Chen, M. L. Structure and functional properties of rice protein-dextran conjugates prepared by the Maillard reaction. International Journal of Food Science and Technology. 2018, 53(2), 372-380. |
| [2] | Liu, J., Ru, Q., Ding, Y. Glycation a promising method for food protein modification: Physicochemical properties and structure, a review. Food Research International. 2012, 49(1), 170-183. |
| [3] | Chobert, J. M., Gaudin, J. C., Dalgalarrondo, M., Haertle, T. Impact of Maillard type glycation on properties of beta-lactoglobulin. Biotechnology Advances. 2006, 24(6), 629-632. |
| [4] | Chen, H. Q., Tu, Z. C., Zhou, Y. R., Xie, Z. H., Zhang, S. Q., Wen, P. W., Liu, J. J., Jiang, Q. N., Wang, H., Hu, Y. M. Insight into the mechanism underlying the reduction of digestibility and IgG/IgE binding ability in ovalbumin during different high-temperature conduction modes-induced glycation. Journal of Agricultural and Food Chemistry. 2024, 72(5), 2801-2812. |
| [5] | Hu, Y. M., Chen, H. Q., Xiao, L., Chu, L. L., Wang, S., Wang, H. Comparison of ovalbumin glycation by microwave irradiation and conventional heating. LWT-Food Science and Technology. 2019, 116, 108560. |
| [6] | Du, P. C., Tu, Z. C., Wang, W., Hu, Y. M. Mechanism of selenium nanoparticles inhibiting advanced glycation end products. Journal of Agricultural and Food Chemistry. 2020, 68, 10586-10595. |
| [7] | Bauer, E. B. Transition metal catalyzed glycosylation reactions-an overview. Organic & Biomolecular Chemistry. 2020, 18(45), 9160-9180. |
| [8] | Kim, E. S., Yaylayan, V. Bis (alaninato) iron (II) complexes as molecular scaffolds for the generation of N, N-di-glycated alanine derivatives in the presence of glucose. Food Chemistry. 2022, 374, 131815. |
| [9] | Chen, B., Huang, W. F., Guo, W. B., Dai, H., Quan, J. H., Zhang, Z. H., Xu, X. Y., Cao, X. D., Zhao, L. Iron-and manganese-catalyzed Maillard chemistry: molecular characteristics, evolution pathways, and implications for soil organic carbon formation. Environmental Science & Technology. 2025, 59(42), 22587-22598. |
| [10] | Hrynets, Y., Bhattacherjee, A., Ndagijimana, M., Martinez, D. J. H., Betti, M. Iron (Fe2+)-catalyzed glucosamine browning at 50 C: identifica-tion and quantification of major flavor compounds for antibacterial activity. Journal of Agricultural and Food Chemistry. 2016, 64(16), 3266-3275. |
| [11] | Xiao, H., Cai, G., Liu, M. Fe2+-catalyzed non-enzymatic glycosylation alters collagen conformation during AGE-collagen formation in vitro. Archives of Biochemistry and Biophysics. 2007, 468(2), 183-192. |
| [12] | Cemil, A. Recent advances and applications in LC-HRMS for food and plant natural products: a critical review. Analytical and Bioanalytical Chemistry. 2020, 412(9), 1973-1991. |
| [13] | Zhang, Q. C., Wang, R., He, J. F., Tang, W., Liu, J. H. Innovative multistep modifications of β-Lactoglobulin for enhanced emulsifying and antioxidant activities. Food Hydrocolloids. 2024, 148, 109465. |
| [14] | Chevalier, F., Chobert, J. M., Popineau, Y., Nicolas, M. G., Haertlé, T. Improvement of functional properties of β-lactoglobulin glycated through the Maillard reaction is related to the nature of the sugar. International Dairy Journal. 2001, 11(3), 145-152. |
| [15] | Leina, E. H., Vanessa, E., Vanessa, C. Maillard reaction: mechanism, influencing parameters, advantages, disad-vantages, and food industrial applications: A Review. Foods. 2025, 14(11), 1881. |
| [16] | Hu, X., Liu, X., Zhang, Y., Yu, Y. L., Zhang, Q. F., Gao, X. Y. Digestive behavior and gut microbiota responses of Glehnia littoralis polysaccharide–Iron complexes: Influence of polysaccharide molecular weight. Food Chemistry: X. 2026, 35, 103777. |
| [17] | Chen, X. X., Zhang, L., Bhesh, B., Zhou, P. Glucose glycation of α-lactalbumin and β-lactoglobulin in glycerol solutions. Journal of Agricultural and Food Chemistry. 2018, 66(40), 10558-10566. |
| [18] | Bian, Z. Y., Tu, Z. C., Wang, H., Hu, Y. M., Liu, G. X. Investigation of the mechanism of 60Co gamma-ray irradiation-stimulated oxidation enhancing the antigenicity of ovalbumin by high-resolution mass spectrometry. Journal of Agricultural and Food Chemistry. 2022, 70(30), 9477-9488. |
| [19] | Hu, Y. M., Guo, H. Z., Wang, H., Yang, Y. F., Tu, Z. C., Huang, T. Insight into the mechanism of urea inhibit ovalbumin-glucose glycation by conventional spectrometry and liquid chromatography-high resolution mass spectrometry. Food Chemistry. 2021, 342, 128340. |
| [20] | Wang, H., Sun, Q., Tan, J. M., Hu, Y. M., Yan, W., Zhen, L., Tu, Z. C. Conformational alteration and the glycated sites in ovalbumin during vacuum freeze-drying induced glycation: A study using conventional spectrometry and liquid chromatography-high resolution mass spectrometry. Food Chemistry. 2020, 318, 126519. |
| [21] | Liu, L., Dong, Q., Kong, Y. M., Kong, Y. R., Yu, Z. Y., Li, B., Yan, H. X., Chen, X., Shen, Y. X. The effect of B-type procyanidin on free radical and metal ion induced β-lactoglobulin glyco-oxidation via mass spectrometry and interaction analysis. Food Research International. 2023, 168, 112744. |
| [22] | Chen, X. M., Kitts, D. D. Correlating changes that occur in chemical properties with the generation of antioxidant capacity in different sugar‐amino acid Maillard reaction models. Journal of Food Science. 2011, 76(6), 831-837. |
| [23] | Qiao, H., Zhu, Z. S., Hua, Y., Guo, X., Huang, M. The role of oxidized myofibrillar protein aggregation and modification in advanced glycation end products formation. Food Chemistry. 2025, 495, 146324. |
| [24] | Murphy, J. M., Powell, B. A., Brumaghim, J. L. Stability constants of bio-relevant, redox-active metals with amino acids: The challenges of weakly binding ligands. Coordination Chemistry Reviews. 2020, 412, 213253. |
| [25] | Sonpasare, K., Lalchandani, D. S., Chenkual, L., Sathala, P. K., Khatoon, R., Porwal, P. K. Effect of glycation-induced concentration-dependent change in albumin structure and alteration in its binding capacity. Journal of Biomolecular Structure and Dynamics. 2025, 43(13), 6488-6497. |
| [26] | Mukherjee, S., Panda, D. Contrasting effects of ferric and ferrous ions on oligomerization and droplet formation of tau: implications in tauopathies and neurodegeneration. ACS Chemical Neuroscience. 2021, 12(23), 4393-4405. |
| [27] | Tu, Z. C., Zhong, B. Z., Wang, H. Identification of glycated sites in ovalbumin under freeze-drying processing by liquid chromatography high-resolution mass spectrometry. Food Chemistry. 2017, 226, 1-7. |
| [28] | She, M. Y., Jia, Z., Zhang, X. Region-selective and site-specific glycation of influenza proteins surrounding the viral envelope membrane. Scientific Reports. 2024, 14(1), 18975. |
| [29] | Chen, Y. J., Liang, L., Liu, X. M., Theodore, P. L., Zhou, P. Effect of fructose and glucose on glycation of β-lactoglobulin in an intermediate-moisture food model system: Analysis by liquid chromatography-mass spectrometry (LC-MS) and data-independent acquisition LC-MS (LC-MSE). Journal of Agricultural and Food Chemistry. 2012, 60(42), 10674-10682. |
| [30] | Doni, D., Passerini, L., Audran, G., Marque, S. R. A., Schulz, M., Santos, J., Costantini, P., Bortolus, M., Carbonera, D. Effects of Fe2+/Fe3+ binding to human frataxin and its D122Y variant, as revealed by site-directed spin labeling (SDSL) EPR complemented by fluorescence and circular dichroism spectroscopies. International Journal of Molecular Sciences. 2020, 21(24), 9619. |
| [31] | Luna, C., Estevez, M. Oxidative damage to food and human serum proteins: Radical-mediated oxidation vs. glyco-oxidation. Food Chemistry. 2018, 267, 111-118. |
APA Style
Wu, X., Jiang, Q., Zhang, X., Zhong, X., Wang, A., et al. (2026). Effects of Fe3+/Fe2+ on Glycation Reaction of β-lactoglobulin. International Journal of Nutrition and Food Sciences, 15(3), 93-103. https://doi.org/10.11648/j.ijnfs.20261503.11
ACS Style
Wu, X.; Jiang, Q.; Zhang, X.; Zhong, X.; Wang, A., et al. Effects of Fe3+/Fe2+ on Glycation Reaction of β-lactoglobulin. Int. J. Nutr. Food Sci. 2026, 15(3), 93-103. doi: 10.11648/j.ijnfs.20261503.11
@article{10.11648/j.ijnfs.20261503.11,
author = {Xiongchen Wu and Qiannan Jiang and Xueying Zhang and Xiangjun Zhong and Amei Wang and Hui Wang and Yueming Hu},
title = {Effects of Fe3+/Fe2+ on Glycation Reaction of
β-lactoglobulin},
journal = {International Journal of Nutrition and Food Sciences},
volume = {15},
number = {3},
pages = {93-103},
doi = {10.11648/j.ijnfs.20261503.11},
url = {https://doi.org/10.11648/j.ijnfs.20261503.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijnfs.20261503.11},
abstract = {Iron ions (Fe2+ and Fe3+) are essential trace elements for the human body, and are often added to various foods, but their effects on protein glycation remain unclear. This study evaluated the differential influences of Fe2+ and Fe3+ on the glycation reaction of β-lactoglobulin (β-Lg)-D-ribose system in terms of glycation degree, protein conformation and the distribution of modification sites. Free amino group contents and HPLC HCD MS/MS analyses indicated that both Fe3+ and Fe2+ could catalyze the glycation process and increase the glycated sites. The system contain Fe2+ exhibited higher glycation degree and more glycation sites (8), and lesser glycation sites were identified in system contain Fe3+ (5) and system without ferric ions (2). Additional sites (L1, K14, K135) were facilitated glycation by Fe2+, and most glycation sites showed higher degree of substitution per peptide (DSP) values when with Fe2+. In comparison with Fe2+, Fe3+ caused more pronounced alterations on both secondary and tertiary protein structure, promoted the β-Lg unfolding, and changed the protein structure to a more unordered form. In conclusion, Fe2+ at a specified concentration was a better choice to promote glycation reaction while maintain the protein structure. This study provide a theoretical basis for protein glycation modification with iron ions at different valence states participated.},
year = {2026}
}
TY - JOUR T1 - Effects of Fe3+/Fe2+ on Glycation Reaction of β-lactoglobulin AU - Xiongchen Wu AU - Qiannan Jiang AU - Xueying Zhang AU - Xiangjun Zhong AU - Amei Wang AU - Hui Wang AU - Yueming Hu Y1 - 2026/05/08 PY - 2026 N1 - https://doi.org/10.11648/j.ijnfs.20261503.11 DO - 10.11648/j.ijnfs.20261503.11 T2 - International Journal of Nutrition and Food Sciences JF - International Journal of Nutrition and Food Sciences JO - International Journal of Nutrition and Food Sciences SP - 93 EP - 103 PB - Science Publishing Group SN - 2327-2716 UR - https://doi.org/10.11648/j.ijnfs.20261503.11 AB - Iron ions (Fe2+ and Fe3+) are essential trace elements for the human body, and are often added to various foods, but their effects on protein glycation remain unclear. This study evaluated the differential influences of Fe2+ and Fe3+ on the glycation reaction of β-lactoglobulin (β-Lg)-D-ribose system in terms of glycation degree, protein conformation and the distribution of modification sites. Free amino group contents and HPLC HCD MS/MS analyses indicated that both Fe3+ and Fe2+ could catalyze the glycation process and increase the glycated sites. The system contain Fe2+ exhibited higher glycation degree and more glycation sites (8), and lesser glycation sites were identified in system contain Fe3+ (5) and system without ferric ions (2). Additional sites (L1, K14, K135) were facilitated glycation by Fe2+, and most glycation sites showed higher degree of substitution per peptide (DSP) values when with Fe2+. In comparison with Fe2+, Fe3+ caused more pronounced alterations on both secondary and tertiary protein structure, promoted the β-Lg unfolding, and changed the protein structure to a more unordered form. In conclusion, Fe2+ at a specified concentration was a better choice to promote glycation reaction while maintain the protein structure. This study provide a theoretical basis for protein glycation modification with iron ions at different valence states participated. VL - 15 IS - 3 ER -
State Key Laboratory of Food Science and Resources, Nanchang University, Nanchang, China; Jiangxi Agricultural Development Group Co., Ltd, Nanchang, China
State Key Laboratory of Food Science and Resources, Nanchang University, Nanchang, China; Nanchang University-Jinggangshan Green Food New Quality Productivity Transformation Center, Nanchang University, Ji’an, China
State Key Laboratory of Food Science and Resources, Nanchang University, Nanchang, China; Nanchang University-Jinggangshan Green Food New Quality Productivity Transformation Center, Nanchang University, Ji’an, China
State Key Laboratory of Food Science and Resources, Nanchang University, Nanchang, China; Nanchang University-Jinggangshan Green Food New Quality Productivity Transformation Center, Nanchang University, Ji’an, China
State Key Laboratory of Food Science and Resources, Nanchang University, Nanchang, China; Nanchang University-Jinggangshan Green Food New Quality Productivity Transformation Center, Nanchang University, Ji’an, China
State Key Laboratory of Food Science and Resources, Nanchang University, Nanchang, China; Nanchang University-Jinggangshan Green Food New Quality Productivity Transformation Center, Nanchang University, Ji’an, China
State Key Laboratory of Food Science and Resources, Nanchang University, Nanchang, China; Nanchang University-Jinggangshan Green Food New Quality Productivity Transformation Center, Nanchang University, Ji’an, China
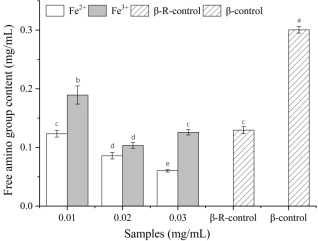
Figure 1. Free amino group contents of β-Lg samples in β-Lg-D-ribose-Fe2+ system and β-Lg-D-ribose-Fe3+ system (0.01-0.03, heat treated-β-Lg-D-ribose system with 0.01-0.03 mg/mL Fe2+/Fe3+ added; β-R-control, heat-treated β-Lg-D-ribose system; β-control, heat-treated β-Lg; Different letters (a–e) denote significant differences among samples (P≤ 0.05)).
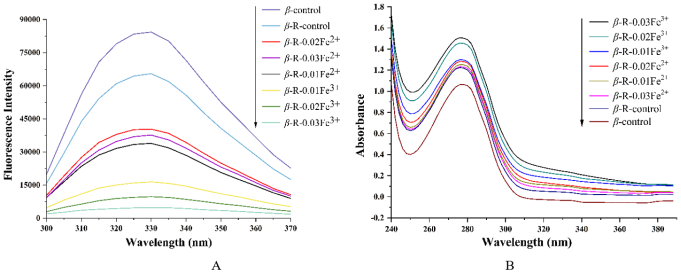
Figure 2. Intrinsic fluorescence spectra (A) and UV absorption spectra (B) of β-Lg samples at different treatments (β-R-0.01-Fe2+, β-R-0.02-Fe2+, β-R-0.03-Fe2+, β-R-0.01-Fe3+, β-R-0.02-Fe3+ and β-R-0.03-Fe3+, heat treated-β-Lg-D-ribose system with 0.01-0.03 mg/mL Fe2+/Fe3+ added; β-R-control, heat-treated β-Lg-D-ribose system; β-control, heat-treated β-Lg).
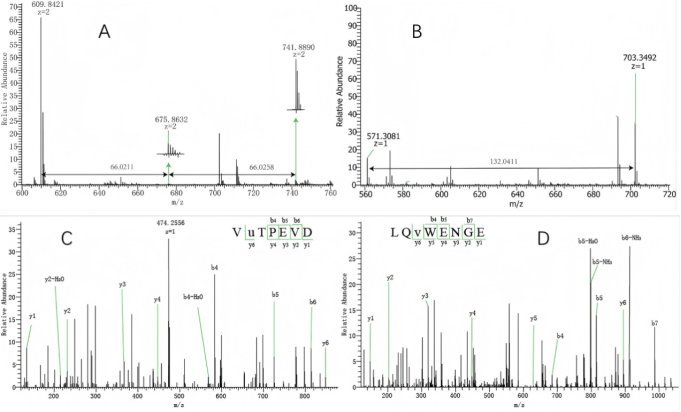
Figure 3. The m/s peaks of glycated peptides by MS spectra at m/z 609.84212+ (A), 571.3081(B). Glycated peptides were identified by mass increasing at 66 Da,132 Da. HCD MS/MS spectrum of mon-glycated peptide 123VuTPEVD129 at arginine with peak m/z at 474.25602+ (C), the glycated site was lysine shown at the glycated peptide 58LQvWENGE65 with peak at 567.27002+ (D).
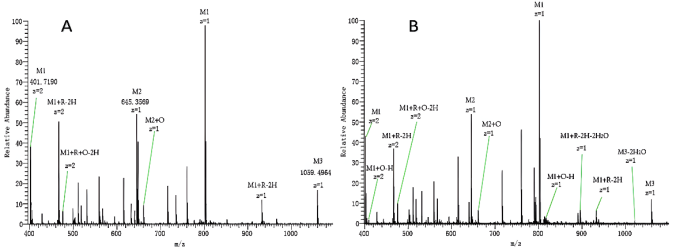
Figure 4. MS spectra with peaks of peptide [M1] 2+, glycated peptide [M1+R-2H] 2+, oxidation glycated peptide [M1+R+O-2H] 2+ when treated with Fe2+ (A). And peptides [M1] 2+, glycated peptide [M1+R-2H] 2+, oxidation peptide [M1+O-H] 2+, oxidation glycated peptide [M1+R+O-2H] 2+, neutral losses glycated peptide [M1+R-2H-2H2O] treated with Fe3+ (B).
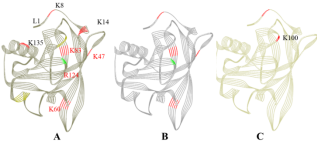
Figure 5. Line ribbon diagram of glycated β-Lg samples (A, β-R-0.03-Fe2+, B, β-R-0.03-Fe3+, C, β-R-control; the glycated lysine residues are colored by red and the glycated arginine residues are colored by green).
Information

