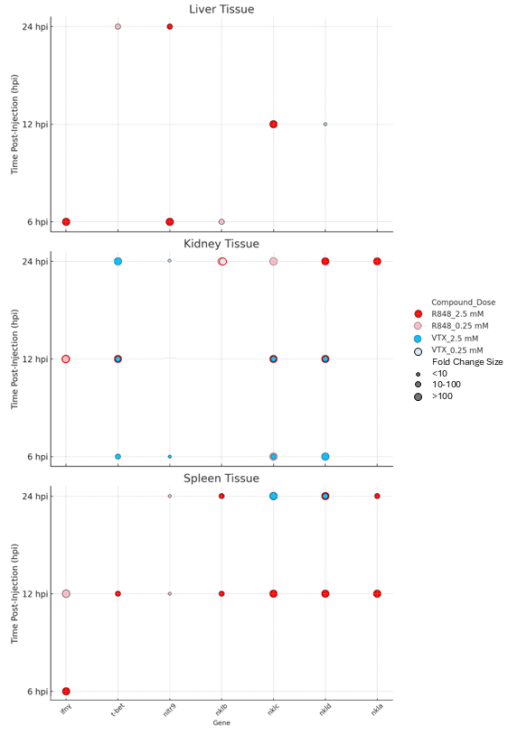
TLR ligands Resiquimod (TLR7/8) and Motolimod (TLR8) are used to activate NK cells and enhance immune defenses. This study reports the differential gene expressions of innate immune markers and Natural Killer (NK) cell lysins in rag1 mutant (rag MT) zebrafish to TLR8 and TLR7/8 activation in liver, kidney, and spleen tissues. Rag MT zebrafish were intracoelomically injected with Motolimod (VTX), Resiquimod (R848), or control saline. Gene expressions of interferon gamma (ifnγ), T-box transcription factor 21 (t-bet), novel immune type receptor 9 (nitr9), and NK lysins a, b, c, and d (nkla, nklb, nklc, nkld) were quantified at 6-, 12-, and 24-hours post-injection using quantitative PCR. We observed that the effects of TLR7/8 and TLR8 stimulation vary depending on the tissue type. R848 significantly upregulated ifnγ, t-bet, nitr9, and NK lysins across various tissues. In contrast, VTX had a more limited effect and primarily influenced nklc and nkld in the kidney and spleen, and nkld in the liver, suggesting tissue-specific responsiveness to TLR8. No significant changes in ifnγ or nkla expression were noted with VTX in any tissues, highlighting the specificity of TLR7 over TLR8 in these responses. Tissue-specific responses revealed dominant activation by TLR7/8 in the liver, particularly influencing ifnγ, t-bet, nitr9, nklb, and nklc. The kidney had high responsiveness to both TLR7/8 and TLR8. The spleen demonstrated broad gene activation by TLR7/8, but only nklc and nkld were significantly upregulated by TLR8. These findings demonstrate that TLR8 has selective effects, while TLR7/8 more broadly activates genes across the liver, kidney, and spleen in rag MT zebrafish. These NK cell gene expression findings suggest exposure to TLR7/8 and TLR8 ligands elicited differential effects across liver, kidney, and spleen tissues of rag MT zebrafish.
| Published in | International Journal of Immunology (Volume 13, Issue 3) |
| DOI | 10.11648/j.iji.20251303.11 |
| Page(s) | 42-51 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Toll Like Receptor Ligands, Natural Killer Cell Activation, Rag Mutant Zebrafish, Tissue-specific Responses
Gene | Oligonucleotide Sequences (5′-3′) | GenBank Accession No. |
|---|---|---|
arp | Fwd: CTGCAAAGATGCCCAGGGA Rev: TTGGAGCCGACATTGTCTGC Probe: [6~FAM] TTCTGAAAATCATCCAACTGCTGGATGACTACC [BHQ1a~ Q] [33] | NM_131580 |
ifnγ | Fwd: CTTTCCAGGCAAGAGTGCAGA Rev: TCAGCTCAAACAAAGCCTTTCG Probe: [6~FAM] AACGCTATGGGCGATCAAGGAAAACGAC[BHQ1a~ Q] [33] | NM_212864 |
t-bet | Fwd: GATCAAGCTCTCTCTGTGATAG Rev: GCTAAAGTCACACAGGTCT Probe: [6~FAM] TTCTGAAGGTCACGGTCACA[BHQ1a~Q] * | NM_001170599.1 |
nitr9 | Fwd: GTCAAAGGGACAAGGCTGATAGTT Rev: GTTCAAAACAGTGCATGTAAGACTCA Probe: [6~FAM] CAAGGTTTGGAAAAGCAC[BHQ1a~Q] | AY570237.1 |
nkla | Fwd: TTTCTGGTCGGCTTGCTCAT Rev: TTCTCATTCACAGCCCGGTC Probe: [6~FAM] TCTGCAGCTCACTGGGAGGTTCGTGA[BHQ1a~Q] | NM_001311794 |
nklb | Fwd: TCCGCAACATCTTTCTGGTCA Rev: AGCCTGCTCATGAATGAAAATGA Probe: [6~FAM] CACGCCTGCAAATCTGAACCACCCA[BHQ1a~Q] | NM_001311792 |
nklc | Fwd: CTGCTTGTGCTGCTCACTTG Rev: AGCACACATGGAGATGAGAACA Probe: [6~FAM] GGGCTTGCAAGTGGGCCATGGGAA[BHQ1a~Q] | NM_001311793.1 |
nkld | Fwd: ACCCTGCTCATCTCCTCTGT Rev: CCCCAGCTAAAGCAAAACCC Probe: [6~FAM] TGCCTGGGATGTGCTGGGCTTGCAA[BHQ1a~Q] | NM_212741.1 |
Treatment | Tissues | Upregulated <10-fold change | Upregulated 10 to 100-fold change | Upregulated >100-fold change | |||
|---|---|---|---|---|---|---|---|
0.25mM | 2.5mM | 0.25mM | 2.5mM | 0.25mM | 2.5mM | ||
R848 | Liver | t-bet (24 hpi*) | nitr9 (24hpi*) nklb (6hpi**) | nitr9 (24 hpi*) | Ifnγ (6hpi**), nitr9 (6hpi**), nklc (12hpi**) | ||
Kidney | t-bet (24 hpi**) | ifnγ (12 hpi**), t-bet (12 hpi**), nkla (24 hpi**), nklc (12 hpi**), nkld (12 hpi**), nkld (24 hpi**) | ifnγ (12 hpi*), nklb (24 hpi*), nklc (6 hpi*), nklc (24 hpi*) | ||||
Spleen | nitr9 (12 hpi*), nitr9 (24 hpi*) | t-bet (12 hpi*), nkla (24 hpi*), nklb (12 hpi*), nklb (24 hpi*), nkld (24 hpi**) | Ifnγ (6 hpi*), Nkla (12 hpi*), Nkld (24 hpi*) | ifnγ (12 hpi**), nklc (12 hpi**), nkld (12 hpi**) | ifnγ (12 hpi*), nklc (24 hpi**), | ||
0.25mM | 2.5mM | 0.25mM | 2.5mM | 0.25mM | 2.5mM | ||
VTX | Liver | Nkld (12 hpi*) | |||||
Kidney | nitr9 (6 hpi*), nitr9 (24 hpi*), | nitr9 (6 hpi*) | Nklb (24 hpi*) | t-bet (6 hpi*), t-bet (12 hpi*), nklc (6 hpi*), nklc (12 hpi*), nkld (6 hpi*), nkld (12 hpi*) | t-bet (24 hpi*), nkld (6 hpi**) | ||
Spleen | Nkld (24 hpi*) | Nklc (24 hpi**) | |||||
NK | Natural Killer |
Rag MT | Rag1 Mutant (rag MT) Zebrafish |
ifnγ | Interferon Gamma |
t-bet | T-box Transcription Factor 21 |
Nitr9 | Novel Immune Type Receptor-9 |
Nkla | NK Lysin a |
Nklb | NK Lysin b |
Nklc | NK Lysin c |
Nkld | NK Lysin d |
NCC | Non-specific Cytotoxic Cell |
LLC | Lymphocyte-like Cell |
PRR | Pathogen Recognition Receptor |
PAMP | Pathogen Associated Molecular Pattern |
R848 | Resiquimod |
VTX | Motolimod VTX2337 |
AMP | Anti-microbial peptides |
MSU IACUC | Mississippi State University Institutional Animal Care and Use Committee |
cDNA | complementary DNA |
PCR | polymerase chain reaction |
Arp | acidic ribosomal phosphoprotein |
ANOVA | Analysis of Variance |
Below is the link to the supplementary material:
| [1] | Sunyer JO. Fishing for mammalian paradigms in the teleost immune system. Vol. 14, Nature Immunology. Nature Publishing Group; 2013. p. 320-6. |
| [2] | DeWitte-Orr S, Edholm ES, Grayfer L. Editorial: Innate Immunity in Aquatic Vertebrates. Vol. 10, Frontiers in Immunology. Frontiers Media S. A.; 2019. |
| [3] | Bailone RL, Fukushima HCS, Ventura Fernandes BH, De Aguiar LK, Corrêa T, Janke H, et al. Zebrafish as an alternative animal model in human and animal vaccination research. Vol. 36, Laboratory Animal Research. BioMed Central Ltd; 2020. |
| [4] | Moss LD, Monette MM, Jaso-Friedmann L, Leary JH, Dougan ST, Krunkosky T, et al. Identification of phagocytic cells, NK-like cytotoxic cell activity and the production of cellular exudates in the coelomic cavity of adult zebrafish. Dev Comp Immunol. 2009; 33(10): 1077-87. |
| [5] | Petrie-Hanson L, Hohn C, Hanson L. Characterization of rag1 mutant zebrafish leukocytes. BMC Immunol. 2009 Feb 3; 10. |
| [6] | Tokunaga Y, Shirouzu M, Sugahara R, Yoshiura Y, Kiryu I, Ototake M, et al. Comprehensive validation of T- and B-cell deficiency in rag1-null zebrafish: Implication for the robust innate defense mechanisms of teleosts. Sci Rep. 2017 Dec 1; 7(1). |
| [7] | Muire PJ, Hanson LA, Wills R, Petrie-Hanson L. Differential gene expression following TLR stimulation in rag1-/-mutant zebrafish tissues and morphological descriptions of lymphocyte-like cell populations. PLoS One [Internet]. 2017; 12(9): 1-22. Available from: |
| [8] | Souza-Fonseca-Guimaraes F, Parlato M, Fitting C, Cavaillon JM, Adib-Conquy M. NK Cell Tolerance to TLR Agonists Mediated by Regulatory T Cells after Polymicrobial Sepsis. The Journal of Immunology. 2012 Jun 15; 188(12): 5850-8. |
| [9] | Rook AH, Gelfand JM, Wysocka M, Troxel AB, Benoit B, Surber C, et al. Topical resiquimod can induce disease regression and enhance T-cell effector functions in cutaneous T-cell lymphoma. Blood. 2015 Sep 17; 126(12): 1452-61. |
| [10] | Chow LQM, Morishima C, Eaton KD, Baik CS, Goulart BH, Anderson LN, et al. Phase Ib trial of the toll-like receptor 8 agonist, motolimod (VTX-2337), combined with cetuximab in patients with recurrent or metastatic SCCHN. Clinical Cancer Research. 2017 May 15; 23(10): 2442-50. |
| [11] | Veneziani I, Alicata C, Pelosi A, Landolina N, Ricci B, D’Oria V, et al. Toll-like receptor 8 agonists improve NK-cell function primarily targeting CD56 bright CD16 - Subset. J Immunother Cancer. 2022 Jan 28; 10(1). |
| [12] | Bender AT, Tzvetkov E, Pereira A, Wu Y, Kasar S, Przetak MM, et al. TLR7 and TLR8 Differentially Activate the IRF and NF-κB Pathways in Specific Cell Types to Promote Inflammation. Immunohorizons. 2020 Feb 1; 4(2): 93-107. |
| [13] | Dietsch GN, Lu H, Yang Y, Morishima C, Chow LQ, Disis ML, et al. Coordinated activation of toll-like receptor8 (TLR8) and NLRP3 by the TLR8 agonist, VTX-2337, ignites tumoricidal natural killer cell activity. PLoS One [Internet]. 2016; 11(2): 1-18. Available from: |
| [14] | Dietsch GN, Randall TD, Gottardo R, Northfelt DW, Ramanathan RK, Cohen PA, et al. Late-stage cancer patients remain highly responsive to immune activation by the selective TLR8 agonist motolimod (VTX-2337). Clinical Cancer Research. 2015 Dec 15; 21(24): 5445-52. |
| [15] | Fischer U, Koppang EO, Nakanishi T. Teleost T and NK cell immunity. Vol. 35, Fish and Shellfish Immunology. Academic Press; 2013. p. 197-206. |
| [16] | Andersson’ M, Gunne H, Agerberth B, Boman A, Bergman T, Olsson’ B, et al. NK-lysin, structure and function of a novel effector molecule of porcine T and NK cells. Vol. 54, Veterinary Immunology and Immunopathology. 1996. |
| [17] | Stinchcombe J, Griffiths G. Secretory Mechanisms in Cell-Mediated Cytotoxicity. Annual Review of Cell and Developmental Biology. 2007; 23: 495-517. |
| [18] | Zhang M, Li MF, Sun L. NKLP27: A teleost NK-lysin peptide that modulates immune response, induces degradation of bacterial DNA, and inhibits bacterial and viral infection. PloS One. (2014) 9(9): e106543-52. |
| [19] | Yu D, Zhao H, Wen Y, Li T, Xia H, Wang Z, et al. Characterization and Functional Evaluation of NK-lysin from Clownfish (Amphiprion ocellaris). Fishes. 2023 Nov 1; 8(11). |
| [20] | Ding FF, Li CH, Chen J. Molecular characterization of the NK-lysin in a teleost fish, Boleophthalmus pectinirostris: Antimicrobial activity and immunomodulatory activity on monocytes/macrophages. Fish Shellfish Immunol. 2019 Sep 1; 92: 256-64. |
| [21] | Acosta J, Roa F, González-Chavarría I, Astuya A, Maura R, Montesino R, et al. In vitro immunomodulatory activities of peptides derived from Salmo salar NK-lysin and cathelicidin in fish cells. Fish Shellfish Immunol. 2019 May 1; 88: 587-94. |
| [22] | Wang GL, Wang MC, Liu YL, Zhang Q, Li CF, Liu PT, et al. Identification, expression analysis, and antibacterial activity of NK-lysin from common carp Cyprinus carpio. Fish Shellfish Immunol. 2018 Feb 1; 73: 11-21. |
| [23] | Zhou QJ, Wang J, Liu M, Qiao Y, Hong WS, Su YQ, et al. Identification, expression and antibacterial activities of an antimicrobial peptide NK-lysin from a marine fish Larimichthys crocea. Fish Shellfish Immunol. 2016 Aug 1; 55: 195-202. |
| [24] | Zhang M, Long H, Sun L. A NK-lysin from Cynoglossus semilaevis enhances antimicrobial defense against bacterial and viral pathogens. Dev Comp Immunol. 2013; 40(3-4): 258-65. |
| [25] | Hirono I, Kondo H, Koyama T, Arma NR, Hwang JY, Nozaki R, et al. Characterization of Japanese flounder (Paralichthys olivaceus) NK-lysin, an antimicrobial peptide. Fish Shellfish Immunol. 2007; 22(5): 567-75. |
| [26] | Wang Q, Wang Y, Xu P, Liu Z. NK-lysin of channel catfish: Gene triplication, sequence variation, and expression analysis. Mol Immunol. 2006 Apr; 43(10): 1676-86. |
| [27] | Pereiro P, Varela M, Diaz-Rosales P, Romero A, Dios S, Figueras A, et al. Zebrafish Nk-lysins: First insights about their cellular and functional diversification. Dev Comp Immunol. 2015 Jul 1; 51(1): 148-59. |
| [28] | Muire PJ, Hanson LA, Petrie-Hanson L. Rapid Natural Killer Cell Gene Responses, Generated by TLR Ligand-Induced Trained Immunity, Provide Protection to Bacterial Infection in rag1−/− Mutant Zebrafish (Danio rerio). Int J Mol Sci. 2025 Feb 1; 26(3). |
| [29] | Klose CSN, Blatz K, d’Hargues Y, Hernandez PP, Kofoed-Nielsen M, Ripka JF, et al. The transcription factor T-bet is induced by IL-15 and thymic agonist selection and controls CD8αα+ intraepithelial lymphocyte development. Immunity. 2014 Aug 21; 41(2): 230-43. |
| [30] | Shah RN, Rodriguez-Nunez I, Eason DD, Haire RN, Bertrand JY, Wittamer V, et al. Development and characterization of anti-Nitr9 antibodies. Adv Hematol. 2012; 2012. |
| [31] | Wei S, JM Z, Chen X, RN S, Liu J, TM O, et al. The zebrafish activating immune receptor Nitr9 signals via Dap12. Immunogenetics. 2007; 59(10): 813-21. |
| [32] | Vivier E, Tomasello E, Baratin M, Walzer T, Ugolini S. Functions of natural killer cells. Vol. 9, Nature Immunology. 2008. p. 503-10. |
| [33] | Vojtech LN, Sanders GE, Conway C, Ostland V, Hansen JD. Host immune response and acute disease in a zebrafish model of Francisella pathogenesis. Infect Immun. 2009 Feb; 77(2): 914-25. |
| [34] | Fitzgerald KA, Kagan JC. Toll-like Receptors and the Control of Immunity. Vol. 180, Cell. Cell Press; 2020. p. 1044-66. |
| [35] | Chang MX. Emerging mechanisms and functions of inflammasome complexes in teleost fish. Vol. 14, Frontiers in Immunology. Frontiers Media S. A.; 2023. |
| [36] | Walker FC, Sridhar PR, Baldridge MT. Differential roles of interferons in innate responses to mucosal viral infections. Vol. 42, Trends in Immunology. Elsevier Ltd; 2021. p. 1009-23. |
| [37] | Murphy K, Weaver C. Janeway’s Immunobiology, 9th edition. 2016. Garland science New York, New York. ISBN: 978-0815345053. |
| [38] | Moore FE, Garcia EG, Lobbardi R, Jain E, Tang Q, Moore JC, et al. Single-cell transcriptional analysis of normal, aberrant, and malignant hematopoiesis in zebrafish. J Exp Med [Internet]. 2016; 213(6): 979-92. Available from: |
APA Style
Muire, P. J., Petrie-Hanson, L. (2025). TLR8 and TLR7/8 Activations Induce Tissue-specific Natural Killer Cell Gene Expressions in Rag1 Mutant Zebrafish Liver, Kidney, and Spleen. International Journal of Immunology, 13(3), 42-51. https://doi.org/10.11648/j.iji.20251303.11
ACS Style
Muire, P. J.; Petrie-Hanson, L. TLR8 and TLR7/8 Activations Induce Tissue-specific Natural Killer Cell Gene Expressions in Rag1 Mutant Zebrafish Liver, Kidney, and Spleen. Int. J. Immunol. 2025, 13(3), 42-51. doi: 10.11648/j.iji.20251303.11
@article{10.11648/j.iji.20251303.11,
author = {Preeti Judith Muire and Lora Petrie-Hanson},
title = {TLR8 and TLR7/8 Activations Induce Tissue-specific Natural Killer Cell Gene Expressions in Rag1 Mutant Zebrafish Liver, Kidney, and Spleen
},
journal = {International Journal of Immunology},
volume = {13},
number = {3},
pages = {42-51},
doi = {10.11648/j.iji.20251303.11},
url = {https://doi.org/10.11648/j.iji.20251303.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.iji.20251303.11},
abstract = {TLR ligands Resiquimod (TLR7/8) and Motolimod (TLR8) are used to activate NK cells and enhance immune defenses. This study reports the differential gene expressions of innate immune markers and Natural Killer (NK) cell lysins in rag1 mutant (rag MT) zebrafish to TLR8 and TLR7/8 activation in liver, kidney, and spleen tissues. Rag MT zebrafish were intracoelomically injected with Motolimod (VTX), Resiquimod (R848), or control saline. Gene expressions of interferon gamma (ifnγ), T-box transcription factor 21 (t-bet), novel immune type receptor 9 (nitr9), and NK lysins a, b, c, and d (nkla, nklb, nklc, nkld) were quantified at 6-, 12-, and 24-hours post-injection using quantitative PCR. We observed that the effects of TLR7/8 and TLR8 stimulation vary depending on the tissue type. R848 significantly upregulated ifnγ, t-bet, nitr9, and NK lysins across various tissues. In contrast, VTX had a more limited effect and primarily influenced nklc and nkld in the kidney and spleen, and nkld in the liver, suggesting tissue-specific responsiveness to TLR8. No significant changes in ifnγ or nkla expression were noted with VTX in any tissues, highlighting the specificity of TLR7 over TLR8 in these responses. Tissue-specific responses revealed dominant activation by TLR7/8 in the liver, particularly influencing ifnγ, t-bet, nitr9, nklb, and nklc. The kidney had high responsiveness to both TLR7/8 and TLR8. The spleen demonstrated broad gene activation by TLR7/8, but only nklc and nkld were significantly upregulated by TLR8. These findings demonstrate that TLR8 has selective effects, while TLR7/8 more broadly activates genes across the liver, kidney, and spleen in rag MT zebrafish. These NK cell gene expression findings suggest exposure to TLR7/8 and TLR8 ligands elicited differential effects across liver, kidney, and spleen tissues of rag MT zebrafish.},
year = {2025}
}
TY - JOUR T1 - TLR8 and TLR7/8 Activations Induce Tissue-specific Natural Killer Cell Gene Expressions in Rag1 Mutant Zebrafish Liver, Kidney, and Spleen AU - Preeti Judith Muire AU - Lora Petrie-Hanson Y1 - 2025/07/23 PY - 2025 N1 - https://doi.org/10.11648/j.iji.20251303.11 DO - 10.11648/j.iji.20251303.11 T2 - International Journal of Immunology JF - International Journal of Immunology JO - International Journal of Immunology SP - 42 EP - 51 PB - Science Publishing Group SN - 2329-1753 UR - https://doi.org/10.11648/j.iji.20251303.11 AB - TLR ligands Resiquimod (TLR7/8) and Motolimod (TLR8) are used to activate NK cells and enhance immune defenses. This study reports the differential gene expressions of innate immune markers and Natural Killer (NK) cell lysins in rag1 mutant (rag MT) zebrafish to TLR8 and TLR7/8 activation in liver, kidney, and spleen tissues. Rag MT zebrafish were intracoelomically injected with Motolimod (VTX), Resiquimod (R848), or control saline. Gene expressions of interferon gamma (ifnγ), T-box transcription factor 21 (t-bet), novel immune type receptor 9 (nitr9), and NK lysins a, b, c, and d (nkla, nklb, nklc, nkld) were quantified at 6-, 12-, and 24-hours post-injection using quantitative PCR. We observed that the effects of TLR7/8 and TLR8 stimulation vary depending on the tissue type. R848 significantly upregulated ifnγ, t-bet, nitr9, and NK lysins across various tissues. In contrast, VTX had a more limited effect and primarily influenced nklc and nkld in the kidney and spleen, and nkld in the liver, suggesting tissue-specific responsiveness to TLR8. No significant changes in ifnγ or nkla expression were noted with VTX in any tissues, highlighting the specificity of TLR7 over TLR8 in these responses. Tissue-specific responses revealed dominant activation by TLR7/8 in the liver, particularly influencing ifnγ, t-bet, nitr9, nklb, and nklc. The kidney had high responsiveness to both TLR7/8 and TLR8. The spleen demonstrated broad gene activation by TLR7/8, but only nklc and nkld were significantly upregulated by TLR8. These findings demonstrate that TLR8 has selective effects, while TLR7/8 more broadly activates genes across the liver, kidney, and spleen in rag MT zebrafish. These NK cell gene expression findings suggest exposure to TLR7/8 and TLR8 ligands elicited differential effects across liver, kidney, and spleen tissues of rag MT zebrafish. VL - 13 IS - 3 ER -
Department of Comparative Biomedical Sciences, College of Veterinary Medicine, Mississippi State University, Starkville, the United States
Department of Comparative Biomedical Sciences, College of Veterinary Medicine, Mississippi State University, Starkville, the United States
Information