1. Introduction
Antimicrobial resistance (AMR) has emerged as one of the most pressing global health challenges of the 21st century. Since the introduction of penicillin in the 1940s, numerous antibacterial agents have been developed and commercialized for therapeutic purposes, significantly reducing morbidity and mortality associated with bacterial infections
| [1] | Muylaert, K., Bastiaens, L., Vandamme, D., & Gouveia, L. (2017). Harvesting of microalgae: Overview of process options and their strengths and drawbacks. Microalgae-based biofuels and bioproducts, 113-132.
https://doi.org/10.1016/B978-0-08-101023-5.00005-4 |
[1]
. However, the widespread and often inappropriate use of antimicrobials has accelerated the emergence and spread of resistant bacteria, threatening to reverse these medical advances. Although progressing more slowly than SARS-CoV-2, AMR represents an equally concerning pandemic
| [2] | Ribeiro, C. F. A., Silveira, G. G. D. O. S., Candido, E. D. S., Cardoso, M. H., Espinola Carvalho, C. M., & Franco, O. L. (2020). Effects of antibiotic treatment on gut microbiota and how to overcome its negative impacts on human health. ACS Infectious Diseases, 6(10), 2544-2559.
https://doi.org/10.1021/acsinfecdis.0c00036 |
[2]
. The World Health Organization recognized AMR as one of the top ten global public health threats, with an estimated 700,000 deaths attributed to drug-resistant infections in 2015
. Without effective interventions, this number could rise to 10 million deaths annually by 2050, surpassing cancer as a leading cause of mortality
.
Globally, approximately 60,000 tons of antibiotics are used annually, with 50% in human medicine and 50% in veterinary applications, including both therapeutic treatments and growth promotion in animal production
| [5] | Parodi, A. L., Verwoerd, D. W., & Bercovich, Z. (2018). Veterinary medicine's increasing relevance to global health and the need for a One Health approach. Revue Scientifique et Technique, 37(1), 21-31.
https://doi.org/10.20506/rst.37.1.2742 |
[5]
. In low- and middle-income countries such as Côte d'Ivoire, antimicrobial misuse is facilitated by self-medication practices, where patients often obtain antimicrobials without prescriptions through unregulated supply chains and insufficient supervision
. This situation accelerates the emergence and spread of drug-resistant pathogens through acquisition of new AMR mechanisms, threatening our ability to treat common infections and increasing healthcare costs by up to €1.5 billion annually
| [7] | Ouedraogo, A. S., Jean Pierre, H., Bañuls, A. L., Ouédraogo, R., & Godreuil, S. (2017). Emergence and spread of antibiotic resistance in West Africa: Contributing factors and threat assessment. Médecine et Santé Tropicales, 27(2), 147-154.
https://doi.org/10.1684/mst.2017.0678 |
[7]
. Furthermore, AMR jeopardizes the achievement of several United Nations Sustainable Development Goals, particularly Goal 3 (Good Health and Well-being).
Livestock production systems represent critical hotspots for AMR development and environmental dissemination. During antimicrobial treatment, bacteria in the gastrointestinal tract rapidly develop or acquire resistance through horizontal transfer of genetic elements such as plasmids, and are subsequently excreted in feces into wastewater streams
. It has been estimated that over 70% of antimicrobials globally used in intensive livestock operations are actually those intended for human use
, making the livestock and meat industry an inadvertent cultivator and distributor of antimicrobial-resistant bacteria (ARB). These resistant organisms enter the food chain and environment through fecal waste, contributing to contamination of surface waters, crops, and meat products, and consequently colonizing wildlife, companion animals, and humans
| [10] | Manyi-Loh, C., Mamphweli, S., Meyer, E., & Okoh, A. (2018). Antibiotic use in agriculture and its consequential resistance in environmental sources: Potential public health implications. Molecules, 23(4), 795.
https://doi.org/10.3390/molecules23040795 |
[10]
.
Antibiotics are only partially metabolized in the animal body, with 30-90% being released into the environment through manure, urine, and feces
| [11] | Daghrir, R., & Drogui, P. (2013). Tetracycline antibiotics in the environment: A review. Environmental Chemistry Letters, 11(3), 209-227. https://doi.org/10.1007/s10311-013-0404-8 |
| [12] | Sharma, B., Dangi, A. K., & Shukla, P. (2016). Contemporary enzyme based technologies for bioremediation: A review. Journal of Environmental Management, 210, 10-22.
https://doi.org/10.1016/j.jenvman.2017.12.075 |
[11, 12]
. This represents a growing concern as their presence can promote selection of antibiotic resistance genes (ARGs), constituting a serious threat to public health
. Among various sources responsible for AMR proliferation, organic solid wastes including municipal waste, agricultural waste, and leachate have been widely reported as potent reservoirs of antibiotic-resistant bacteria and ARGs
. Consequently, manure management practices play a crucial role in controlling AMR dissemination in agricultural environments, particularly when these materials are used as fertilizers with or without additional treatment.
In recent decades, anaerobic digestion (AD) has gained recognition as an environmentally sound and efficient technology for treating organic wastes. AD contributes to waste reduction through resource recovery, recycling, and biological treatment
| [14] | Okeke, E. S., Ita, R. E., Okoye, C. O., Ezeorba, T. P. C., & Mao, G. (2020). A critical review of life cycle assessment studies of renewable diesel fuels. Energies, 13(19), 5137.
https://doi.org/10.3390/en13195137 |
[14]
, while simultaneously producing renewable energy in the form of biogas. The process operates under oxygen-free conditions where complex organic matter is broken down by microbial consortia through sequential stages: hydrolysis, acidogenesis, acetogenesis, and methanogenesis
. Beyond energy production, AD has been proposed as a method to reduce pathogen loads and potentially mitigate AMR dissemination from livestock waste.
However, knowledge regarding the persistence of human pathogens and AMR throughout the manure treatment chain remains limited. The effectiveness of AD in eliminating resistant bacteria depends on multiple factors, including microbial characteristics, operational conditions such as temperature and hydraulic retention time (HRT), and the initial contamination level
| [16] | Smith, S. R., Lang, N. L., Cheung, K. H. M., & Spanoudaki, K. (2005). Factors controlling pathogen destruction during anaerobic digestion of biowastes. Waste Management, 25(4), 417-425. https://doi.org/10.1016/j.wasman.2005.02.010 |
| [17] | Pandey, P. K., & Soupir, M. L. (2011). Escherichia coli inactivation kinetics in dairy manure under aerobic and anaerobic conditions. Bioresource Technology, 107, 204-211.
https://doi.org/10.1016/j.biortech.2011.12.017 |
[16, 17]
. Previous studies have shown variable results: while mesophilic digestion (25-40°C) achieves substantial reductions in indicator organisms, certain bacterial groups particularly spore-forming species demonstrate remarkable persistence
| [18] | Bagge, E., Sahlström, L., & Albihn, A. (2005). The effect of hygienic treatment on the microbial flora of biowaste at biogas plants. Water Research, 39(20), 4879-4886.
https://doi.org/10.1016/j.watres.2005.03.016 |
| [19] | Sahlström, L. (2003). A review of survival of pathogenic bacteria in organic waste used in biogas plants. Bioresource Technology, 87(2), 161-166.
https://doi.org/10.1016/S0960-8524(02)00168-2 |
[18, 19]
. Furthermore, regulatory frameworks governing agricultural use of digestates typically focus on pathogen reduction but may not adequately address AMR concerns
| [20] | Venglovsky, J., Sasakova, N., & Placha, I. (2009). Pathogens and antibiotic residues in animal manures and hygienic and ecological risks related to subsequent land application. Bioresource Technology, 100(22), 5386-5391.
https://doi.org/10.1016/j.biortech.2009.03.068 |
| [21] | Martens, W., & Böhm, R. (2009). Overview of the ability of different treatment methods for liquid and solid manure to inactivate pathogens. Bioresource Technology, 100(22), 5374-5378. https://doi.org/10.1016/j.biortech.2009.01.014 |
[20, 21]
.
The emergence and abundance of antimicrobial-resistant strains raise critical questions about whether waste treatment and valorization methods adequately reduce or eliminate dissemination of these micropollutants. This concern is particularly relevant within the One Health framework, which recognizes AMR as a shared threat affecting human, animal, and environmental health
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[22]
. In the context of West Africa, where intensive livestock production is expanding and antimicrobial stewardship programs remain limited, understanding the fate of resistant bacteria during manure processing is essential for developing evidence-based waste management policies.
Despite growing interest in AD as a sustainable waste management solution, comprehensive data on the survival and resistance profiles of pathogenic bacteria throughout the digestion process in tropical conditions remain scarce. Most previous studies have been conducted in temperate regions with controlled temperature systems, limiting their applicability to resource-constrained settings where ambient-temperature systems predominate. Additionally, while conventional culture-based methods provide valuable information, modern molecular identification techniques such as Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (MALDI-TOF MS) offer rapid, accurate species-level identification that can enhance our understanding of microbial community dynamics during AD.
This study aimed to address these knowledge gaps by investigating the fate of potentially pathogenic bacteria and their antimicrobial resistance profiles during mesophilic anaerobic digestion of pig slurry under ambient tropical conditions in Abidjan, Côte d'Ivoire. The specific objectives were to: (i) evaluate the impact of anaerobic digestion on bacterial community composition and pathogen loads over a 56-day retention period; (ii) identify bacterial isolates using MALDI-TOF MS technology; and (iii) characterize phenotypic antibiotic resistance profiles of pathogenic isolates throughout the AD process. The findings will contribute to evidence-based recommendations for safe agricultural valorization of digestates in the context of AMR mitigation strategies.
2. Materials and Methods
2.1. Study Sites and Manure Collection
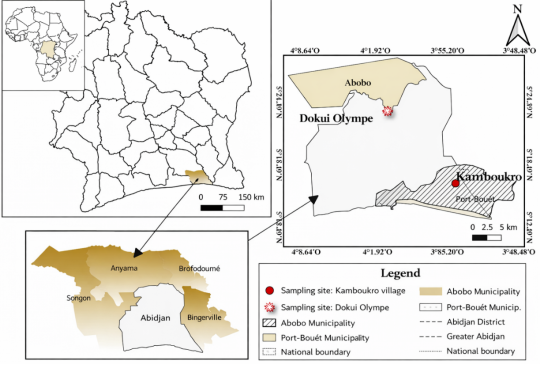
This study was conducted between January and June 2022 in two-pig farming facilities located in the communes of Abobo (Dokui Olymphe district) and Port-Bouët (Kamboukro district) in Abidjan, Côte d'Ivoire (
Figure 1). These sites were selected based on their intensive pig production systems and accessibility for regular sampling throughout the experimental period. Fresh pig manure was collected directly from the farm floors immediately after excretion to minimize environmental contamination and oxidation. Prior to use, all collected manure was manually cleaned to remove bones, bedding materials, and other debris that could interfere with the anaerobic digestion process. A total of 320 kg of cleaned fresh manure was collected from each site and transported in sealed containers to prevent spillage and odor emission during transfer.
Figure 1. Map showing the location of the various sites.
2.2. Biodigester Design and Operational Conditions
Two identical pilot-scale continuous anaerobic digesters were installed, one at each farm location. The digestors utilized the Puxin biogas system (Puxin Technology Co., China), specifically designed for small-scale agricultural applications in tropical climates. Each unit consisted of a flexible digestion bag with a total volume of 1600 L manufactured from specialized material resistant to aging, acids, alkalis, and ultraviolet radiation. The system incorporated an integrated gas storage bag, a support frame for structural stability, a feeding pump for substrate introduction, and a discharge outlet for effluent removal. Additional components included a biogas filter for gas purification and a pressure pump to supply gas to appliances (
Figure 2). The digestors were operated as batch systems with a single initial loading and a hydraulic retention time of 56 days. For initial feeding, 320 kg of fresh pig manure was mixed with 1280 L of water to achieve approximately 25% total solids content, facilitating homogenization and flow within the digestion bag. The mixture was introduced until the digestor reached one-quarter of its total volume, allowing adequate headspace for biogas accumulation. Both units operated under ambient temperature conditions ranging from 25°C to 36.9°C, characteristic of the mesophilic range in tropical West African environments. No external heating was applied, making the system representative of resource-limited agricultural settings. The digestor at Port-Bouët was installed on-site at the pig farm, while the Abobo unit was installed at Université Nangui Abrogoua for closer monitoring accessibility.
Figure 2. Puxin biogas system.
2.3. Sampling Strategy and Monitoring
Throughout the 56-day digestion period, process performance was monitored through regular measurement of key operational parameters. Temperature was recorded daily using a digital thermometer inserted into the digestion medium. The pH was measured weekly using a portable pH meter (Hanna Instruments, USA) calibrated with standard buffer solutions. Biogas production was quantified daily using a water displacement method, and methane content was determined weekly by gas chromatography following standard methods
| [23] | American Public Health Association. (2017). Standard methods for the examination of water and wastewater (23rd ed.). APHA Press. |
[23]
. Total solids and volatile solids were measured at the beginning and end of the digestion period according to standard procedures
| [24] | AOAC International. (2019). Official methods of analysis of AOAC International (21st ed.). AOAC International. |
[24]
. For microbiological analysis, 24 samples were collected across both digestors over the experimental period. Sampling was conducted at four specific time points to track microbial community dynamics: Day 0 (initial influent, n=6 samples total from both sites), Day 14 (n=6), Day 28 (n=6), and Day 56 (n=6). At each sampling point, triplicate 20 mL samples were aseptically collected from each digestor using sterile bottles that were immediately sealed and transported on ice to the Laboratory of Biotechnology and Food Microbiology at Université Nangui ABROGOUA. All samples were processed within one hour of collection to minimize changes in microbial viability during storage.
2.4. Microbiological Analyses
2.4.1. Bacterial Enumeration
Bacterial enumeration targeted key indicator organisms and potential pathogens relevant to assessing the sanitary quality of digestate for agricultural use. For each sample, decimal serial dilutions were prepared from 10-¹ to 10⁻⁸ in sterile buffered peptone water (BPW, Condalab, France). Aliquots of 0.1 mL from appropriate dilutions were spread-plated onto selective media in duplicate. The lowest dilution yielding between 30 and 300 colonies was used for enumeration, and counts were expressed as colony-forming units per gram of initial material (CFU g⁻¹). Gram-positive cocci were enumerated on specific media: Enterococcus spp. on Bile Esculin Agar (Himedia Laboratories, India) and Staphylococcus spp. on Baird-Parker Agar supplemented with egg yolk tellurite (Condalab, France). Clostridium difficile was cultured on Clostridium Difficile Agar (Condalab, France). All Gram-positive bacteria were incubated at 37°C for 24 hours under aerobic conditions, except for Clostridium which required anaerobic incubation in anaerobic jars with gas-generating sachets. Gram-negative bacteria were cultured using selective enrichment and plating protocols. Salmonella spp. were first enriched in Rappaport-Vassiliadis broth at 42°C for 24 hours, then streaked onto Salmonella-Shigella (SS) Agar and incubated at 37°C for 24 hours. Yersinia spp. were plated on Cefsulodin- Irgasan-Novobiocin (CIN) Agar, and Pseudomonas spp. on King A and King B Agar. Enterobacteriaceae were cultured on Eosin Methylene Blue (EMB) Agar (Himedia Laboratories, India), with lactose-fermenting colonies appearing as metallic green. All Gram-negative bacteria were incubated aerobically at 37°C for 24 hours. Colony counts from duplicate plates were averaged, and the number of CFU per gram of initial sample was calculated using the following formula:
N = ΣC / [(n₁ + 0.1n₂) × d]
where N represents the number of CFU per gram of initial product, ΣC is the sum of colonies counted on all retained plates, n₁ is the number of plates counted at the first retained dilution, n₂ is the number of plates counted at the second retained dilution, and d is the dilution factor of the first retained dilution.
2.4.2. Isolation and Presumptive Identification
From each selective medium showing growth, three to five morphologically representative colonies were selected and subcultured in nutrient broth (Conda Lab, France) at 37°C for 24 hours. Pure cultures were preserved by freezing at -80°C in nutrient broth containing 20% glycerol for subsequent identification and antimicrobial susceptibility testing. Presumptive identification was performed using conventional microbiological methods. Gram-positive cocci were characterized by Gram staining, catalase testing, and DNase production for staphylococci, while esculin hydrolysis was used for enterococci. Gram-negative bacteria were characterized by Gram staining, oxidase testing, motility testing, and carbohydrate fermentation patterns (glucose, lactose, and sucrose) following established protocols described by Le Minor and colleagues
| [25] | Grimont, P. A. D., & Weill, F. X. (2007). Antigenic formulae of the Salmonella serovars (9th ed.). WHO Collaborating Centre for Reference and Research on Salmonella, Institut Pasteur. |
[25]
.
2.4.3. MALDI-TOF MS Identification
Definitive species-level identification was achieved using Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry on a VITEK MS system (bioMérieux, France). For each isolate, one to two colonies from fresh overnight cultures were directly deposited onto the manufacturer's proprietary sample plates following standard protocols. After air-drying, 1 μL of CHCA matrix solution (α-cyano-4-hydroxycinnamic acid, bioMérieux) was applied to each spot and allowed to crystallize at room temperature. The VITEK MS instrument utilized a fixed-focus 337 nm nitrogen laser operating at 50 Hz frequency. Mass spectra were acquired in the range of 2,000 to 20,000 Da, generating a unique composite mass spectrum for each isolate. Escherichia coli ATCC 8739 was used as a positive control for quality assurance. Spectral analysis was performed using VITEK MS IVD software version 3.2 with the manufacturer's comprehensive clinical database for bacterial and fungal identification. Identification confidence scores were interpreted as follows: 60-100% probability indicated excellent discrimination and reliable identification at the species level, scores below 60% represented low discrimination with multiple possible identifications, and no identification was assigned when spectral quality was insufficient or no database match was found.
2.5. Antimicrobial Susceptibility Testing
Phenotypic antibiotic resistance profiles were determined for all confirmed pathogenic isolates using the disk diffusion method on Mueller-Hinton agar (Himedia Laboratories, India) according to guidelines established by the Comité de l'Antibiogramme de la Société Française de Microbiologie (CASFM) version 2023
| [26] | Antibiogram Committee of the French Society for Microbioogy. (2023). 2023 Recommendations [CA-SFM guidelines 2023]. French Society for Microbiology.
https://www.sfm-microbiologie.org/ |
[26]
. Bacterial suspensions were prepared from fresh overnight cultures and adjusted to an optical density of 0.08-0.10 at 625 nm, corresponding to approximately 1 × 10⁸ CFU mL
⁻¹. The agar surface was inoculated by flooding with the bacterial suspension, and excess liquid was removed. After drying for 15 minutes, antibiotic disks were applied using sterile forceps. The antibiotic panel was selected based on veterinary antibiotics commonly used in pig production in Côte d'Ivoire and therapeutic agents relevant to human health (
Table 1). Different antibiotic combinations were tested depending on bacterial groups to match clinical relevance and expected resistance mechanisms. Following 18-24 hours of incubation at 37°C, inhibition zone diameters were measured to the nearest millimeter using a digital caliper. Results were interpreted as susceptible, intermediate, or resistant by comparing measured diameters to critical breakpoints published in CASFM 2023 guidelines. The Multiple Antibiotic Resistance (MAR) index was calculated for each bacterial group to assess the extent of multidrug resistance. The MAR index was computed as the ratio of the number of antibiotics to which an isolate showed resistance divided by the total number of antibiotics tested against that isolate
| [27] | Krumperman, P. H. (1983). Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Applied and Environmental Microbiology, 46(1), 165-170.
https://doi.org/10.1128/aem.46.1.165-170.1983 |
[27]
. A MAR index greater than 0.2 indicates high-risk sources of contamination where antibiotics are frequently used.
Table 1. Antimicrobial Agents Tested by Bacterial Group.
Bacterial Group | Antibiotics Tested (Class) |
Bacillus spp. | MEM (carbapenem), CIP, NOR (fluoroquinolones), VA (glycopeptide) |
Clostridium spp. | A, AUG (β-lactams), DA (lincosamide), C (phenicol), IMI (carbapenem), TGC (glycylcycline) |
Enterococcus spp. | FEP (cephalosporin), ATM (monobactam), LEV (fluoroquinolone), TGC (glycylcycline), AK (aminoglycoside) |
Enterobacteriaceae | MEM (carbapenem), CAZ (cephalosporin), A, TIC (penicillins), TC (tetracycline), CIP (fluoroquinolone) |
Salmonella spp. | LEV, CIP (fluoroquinolones), AK (aminoglycoside), CAZ, CRO (cephalosporins) |
Pseudomonas spp. | CPM (cephalosporin), IMI (carbapenem), CIP (fluoroquinolone), TC (tetracycline), AK (aminoglycoside) |
Staphylococcus spp. | DA (lincosamide), C (phenicol), K (aminoglycoside), E (macrolide), FA (fusidic acid) |
Yersinia spp. | A, PIP, TIC (penicillins), TOB, AK (aminoglycosides), ETP, MEM (carbapenems) |
2.6. Statistical Analyses
All microbiological counts were log₁₀ transformed prior to statistical analysis to achieve normal distribution. One-way analysis of variance (ANOVA) followed by Tukey's post hoc test for multiple comparisons was performed to compare bacterial loads across sampling times and between digestors. Student's t-test was used to compare reduction rates between the two sites. Statistical significance was set at p < 0.05. All analyses were conducted using GraphPad Prism version 9.0 (GraphPad Software, USA).
3. Results
3.1. Biodigester Performance and Operational Conditions
Both pilot-scale biodigesters operated successfully throughout the 56-day experimental period under mesophilic conditions (
Table 2). The initial pH of the pig slurry ranged from 6.50 ± 0,05 to 6.88 ± 0.03, increasing progressively to 8.40 ± 0,05 - 9.40 ± 0,06 by the end of the digestion period. This pH elevation reflects efficient consumption of volatile fatty acids produced during acidogenesis and active methanogenesis
, although it may also indicate some accumulation of free ammonia
| [16] | Smith, S. R., Lang, N. L., Cheung, K. H. M., & Spanoudaki, K. (2005). Factors controlling pathogen destruction during anaerobic digestion of biowastes. Waste Management, 25(4), 417-425. https://doi.org/10.1016/j.wasman.2005.02.010 |
[16]
. Ambient temperature varied between 25.0°C and 37.9°C throughout the study period, characteristic of the tropical mesophilic range in Côte d'Ivoire.
Process performance indicators demonstrated effective organic matter degradation, with total solids reduction reaching 64% and volatile solids reduction achieving 70%. Daily biogas production ranged from 290 to 400 L, with methane content consistently maintained between 65% and 76%, indicating stable methanogenic activity
. No significant differences in methane composition were observed between the two digestors (p = 0.5), suggesting similar microbial community dynamics and process stability at both sites.
Table 2. Biodigester performance parameters.
Parameters | Abobo | Port-Bouët |
Initial pH (Day 0) | 6.50 ± 0,03 a | 6.88 ± 0.03 a |
Final pH (Day 56) | 8.40± 0,05 a | 9.40± 0,06 b |
Temperature range (°C) | 25.0 ± 0.09 a – 37.03± 0.001 b | 25.4± 0.02 a - 37.9±0.006 b |
Total solids reduction (%) | 25,51 ± 1,52b | 29,51 ± 1,13a |
Volatile solids reduction (%) | 56 | 55 |
Biogas production (m³·day⁻¹) Average biogas production during the active phase (D0–D28) | 0.29 ± 0.001 a | 0.40 ±0.003 b |
Methane content (%) | 80 | 82 |
Hydraulic retention time (days) | 56 | 56 |
3.2. Evolution of Bacterial Loads During Anaerobic Digestion
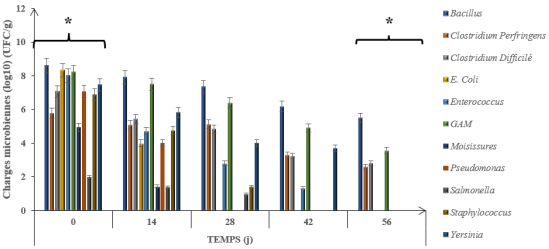
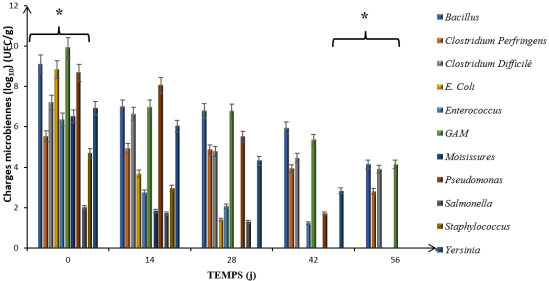
Bacterial enumeration revealed substantial reductions in most microbial groups over the 56-day digestion period, although patterns varied considerably among different bacterial groups (
Figure 3 and
Figure 4). At both sites, initial bacterial loads were high, with most groups ranging from 10⁵ to 10⁹ CFU g⁻¹ at Day 0.
For
Escherichia coli, initial loads of 8.33 log₁₀ CFU g⁻¹ (Abobo) and 8.84 log₁₀ CFU g⁻¹ (Port-Bouët) decreased rapidly, with the bacterium becoming undetectable by Day 56 at both sites. Similarly,
Enterococcus spp., starting at 8.01 log₁₀ CFU g⁻¹ (Abobo) and 6.36 log₁₀ CFU g⁻¹ (Port-Bouët), showed progressive decline and complete elimination by the end of the process
| [17] | Pandey, P. K., & Soupir, M. L. (2011). Escherichia coli inactivation kinetics in dairy manure under aerobic and anaerobic conditions. Bioresource Technology, 107, 204-211.
https://doi.org/10.1016/j.biortech.2011.12.017 |
| [18] | Bagge, E., Sahlström, L., & Albihn, A. (2005). The effect of hygienic treatment on the microbial flora of biowaste at biogas plants. Water Research, 39(20), 4879-4886.
https://doi.org/10.1016/j.watres.2005.03.016 |
[17, 18]
.
Pseudomonas spp.,
Salmonella spp.,
Staphylococcus spp., and
Yersinia spp. followed similar patterns of complete inactivation by Day 56.
In contrast, spore-forming bacteria demonstrated significantly greater persistence
.
Bacillus spp. exhibited moderate reductions of 5.49 log₁₀ CFU g⁻¹ at Abobo and 4.13 log₁₀ CFU g⁻¹ at Port-Bouët but remained detectable at 10⁵ and 10⁴ CFU g⁻¹, respectively. C
lostridium perfringens showed reductions of 2.61 log₁₀ CFU g⁻¹ (Abobo) and 2.79 log₁₀ CFU g⁻¹ (Port-Bouët), while
Clostridium difficile achieved 2.82 log₁₀ CFU g⁻¹ (Abobo) and 3.89 log₁₀ CFU g⁻¹ (Port-Bouët) reductions. Despite these substantial decreases, all spore-forming bacteria remained detectable at the conclusion of the digestion period.
Mesophilic aerobic bacteria (GAM), used as general indicators of process hygienization, demonstrated excellent reductions of 3.56 log₁₀ CFU g⁻¹ at Abobo and 4.14 log₁₀ CFU g⁻¹ at Port-Bouët, representing the highest logarithmic reductions observed in this study. Statistical analysis revealed that bacterial load reductions differed significantly across sampling times (p < 0.05), with the most substantial decreases occurring between Day 14 and Day 28.
Figure 3. Evolution of microbial loads during anaerobic digestion - Abobo site.
Figure 4. Evolution of microbial loads during anaerobic digestion - Port-bouët site.
3.3. Bacterial Identification by MALDI-TOF MS
From the 245 bacterial isolates subjected to MALDI-TOF MS analysis, 103 isolates (42.0%) from both sites were definitively identified and selected for further characterization (
Table 3). The identification success rate was 93.4%, with 76.2% identified at the species level and 17.2% at the genus level. Failed or ambiguous identifications, primarily attributed to incomplete database entries or insufficient spectral quality were evaluated to 10.9%. Isolates were distributed across eight bacterial genera spanning both
Firmicutes and
Proteobacteria phyla. The temporal distribution of isolates from the Abobo site revealed distinct patterns of bacterial persistence throughout the digestion process. At Day 0, all eight genera were represented with five isolates each (n = 40 total). This diversity was maintained through Day 14 (n = 40), but by Day 28,
Escherichia and
Pseudomonas were no longer detected (n = 30 total). By Day 56, only
Bacillus and
Clostridium difficile persisted (n = 10 total). Among the 103 confirmed isolates from both sites,
Firmicutes accounted for 54 isolates (52.4%), including spore-forming genera (
Bacillus,
Clostridium) and Gram-positive cocci (
Enterococcus,
Staphylococcus).
Proteobacteria comprised 49 isolates (47.6%), consisting entirely of Gram-negative rods (
Escherichia,
Salmonella,
Yersinia,
Pseudomonas).
Table 3. Distribution of Bacterial Isolates by MALDI-TOF MS.
Phylum | Genus | Bacterial species identified by MALDI-TOF | N° of isolates identified | Homology rate (%) |
| Bacillus | Bacillus cereus | 5 | 99.9% |
| n=16 | Bacillus licheniformis | 7 | 99.9% |
Firmicutes | | Lysinibacillus fusiformis | 4 | 99.9% |
n=54 | Clostridium | Clostridium perfringens | 6 | 99.9% |
| n=10 | Clostridioides difficile | 4 | 99.9% |
| Staphylococcus n=11 | Staphylococcus aureus | 11 | 99.9% |
| Enterococcus | Enterococus faecium | 9 | 99.9% |
| n=17 | Enterococus faecalis | 8 | 99.9% |
| Escherichia n=11 | Escherichia coli | 11 | 99.9% |
| Pseudomonas | Pseudomonas aeruginosa | 2 | 99.9% |
Proteobacteria | n=09 | Pseudomonas spp | 7 | 99.9% |
n=49 | Salmonella n=11 | Salmonella enterica | 11 | 99.9% |
| Yersinia n=18 | Yersinia enterocolitica | 18 | 99.9% |
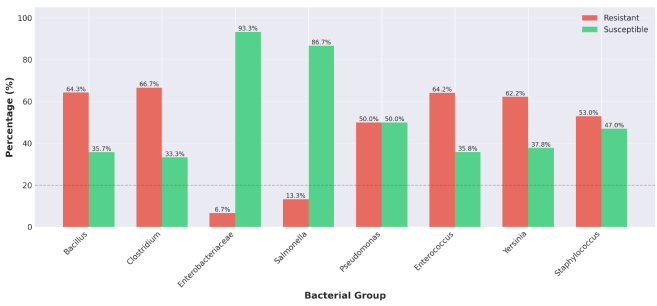
3.4. Antimicrobial Susceptibility Profiles
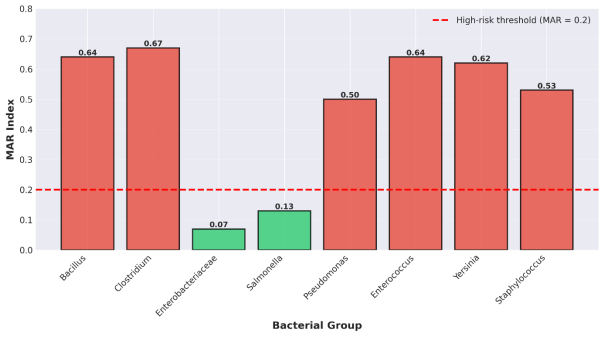
Antimicrobial susceptibility testing revealed substantial variation in resistance patterns among bacterial groups (
Table 5,
Figure 5). The antibiotic panel tested included representatives from major classes, β-lactams, fluoroquinolones, aminoglycosides, glycopeptides, tetracyclines, and other clinically relevant agents.
3.4.1. Gram-positive Spore-Forming Bacteria
Bacillus spp. (n = 4 isolates) demonstrated high resistance levels, with a mean resistance rate of 64.3% across tested antibiotics. Resistance was particularly pronounced against fluoroquinolones (ciprofloxacin, norfloxacin) and carbapenems (meropenem), with 57.1% of isolates showing resistant phenotypes. The MAR index for Bacillus spp. was 0.64, substantially exceeding the threshold of 0.2 that indicates high-risk contamination sources. Clostridium spp. (n = 6 isolates) exhibited even higher resistance levels (66.7% mean resistance, MAR index 0.67). While all isolates remained susceptible to chloramphenicol (100% susceptibility), resistance to β-lactams was common, with 80% of isolates resistant to imipenem and variable resistance to ampicillin (30%) and amoxicillin-clavulanic acid (20%).
3.4.2. Gram-positive Cocci
Enterococcus spp. (n = 5 isolates) displayed a mean resistance rate of 64.2% (MAR index 0.64). Resistance was widespread across multiple antibiotic classes: 88% of isolates were resistant to levofloxacin, 90% to tigecycline, and 90% to amikacin. Staphylococcus spp. (n = 11 isolates) presented a mean resistance rate of 53.0% (MAR index 0.53), with moderate resistance to lincosamides (clindamycin, 62.8%) and variable resistance to aminoglycosides, macrolides, and phenicols.
3.4.3. Gram-negative Bacteria
Enterobacteriaceae (n = 6 isolates) demonstrated remarkably low resistance rates (mean 6.7%, MAR index 0.07), representing the only bacterial group with MAR index below 0.2. Susceptibility exceeded 93% for all tested antibiotics, including carbapenems, cephalosporins, fluoroquinolones, and aminoglycosides. Salmonella spp. (n = 5 isolates) similarly showed low resistance (mean 13.3%, MAR index 0.13), with 100% susceptibility to levofloxacin, amikacin, ceftazidime, and ceftriaxone. Only ciprofloxacin showed elevated resistance (66.7%). Pseudomonas spp. (n = 5 isolates) exhibited intermediate resistance levels (mean 50.0%, MAR index 0.50), reflecting intrinsic resistance mechanisms including efflux pumps and low outer membrane permeability. Yersinia spp. (n = 6 isolates) demonstrated high resistance (mean 62.2%, MAR index 0.62), with particularly elevated resistance to ampicillin (96.3%), piperacillin (86.0%), and ticarcillin (79.3%). Resistance decreased for tobramycin (67.0%), amikacin (27.5%), and ertapenem (17.3%).
Figure 5. Antibiotic Resistance and Susceptibility Profiles.
Table 4. Antibiotic Resistance Profiles and MAR Indices.
Bacterial Group | n | Resistance (%) | Susceptibility (%) | MAR Index | Risk |
Bacillus spp. | 4 | 64.3 | 35.7 | 0.64 | High |
Clostridium spp. | 6 | 66.7 | 33.3 | 0.67 | High |
Enterobacteriaceae | 6 | 6.7 | 93.3 | 0.07 | Low |
Salmonella spp. | 5 | 13.3 | 86.7 | 0.13 | Low |
Pseudomonas spp. | 5 | 50.0 | 50.0 | 0.50 | High |
Enterococcus spp. | 5 | 64.2 | 35.8 | 0.64 | High |
Yersinia spp. | 6 | 62.2 | 37.8 | 0.62 | High |
Staphylococcus spp. | 11 | 53.0 | 47.0 | 0.53 | High |
3.5. Multiple Antibiotic Resistance (MAR) Indices
Analysis of MAR indices revealed that six of the eight bacterial groups (75%) exceeded the critical threshold of 0.2 (
Figure 6).
Clostridium spp. exhibited the highest MAR index (0.67), followed by Bacillus spp. and
Enterococcus spp. (both 0.64), Yersinia spp. (0.62),
Staphylococcus spp. (0.53), and
Pseudomonas spp. (0.50). In contrast, Enterobacteriaceae (MAR 0.07) and
Salmonella spp. (MAR 0.13) remained below the 0.2 threshold, suggesting these isolates originated from environments with lower antimicrobial selective pressure. The mean MAR index across all groups was 0.47. Statistical analysis revealed no significant correlation between bacterial persistence through anaerobic digestion and MAR index (r = 0.23, p = 0.58), indicating that antibiotic resistance did not confer survival advantages under anaerobic digestion conditions.
Figure 6. Multiple Antibiotic Resistance (MAR) Indices by Bacterial Group.
4. Discussion
The present study demonstrates that mesophilic anaerobic digestion achieves substantial reductions in bacterial pathogens from pig slurry, with logarithmic reductions ranging from 2.7 to 5.8 log
10 CFU g
-1 depending on bacterial group and site. However, our results reveal important limitations in the complete elimination of spore-forming bacteria, which has critical implications for digestate safety and agricultural reuse. The complete elimination of
Escherichia coli,
Enterococcus spp., and
Salmonella spp. by Day 56 aligns well with regulatory expectations for digestate hygienization
| [16] | Smith, S. R., Lang, N. L., Cheung, K. H. M., & Spanoudaki, K. (2005). Factors controlling pathogen destruction during anaerobic digestion of biowastes. Waste Management, 25(4), 417-425. https://doi.org/10.1016/j.wasman.2005.02.010 |
| [21] | Martens, W., & Böhm, R. (2009). Overview of the ability of different treatment methods for liquid and solid manure to inactivate pathogens. Bioresource Technology, 100(22), 5374-5378. https://doi.org/10.1016/j.biortech.2009.01.014 |
[16, 21]
. Sahlström
reported similar patterns, demonstrating that non-spore-forming pathogens are generally inactivated during mesophilic AD at retention times exceeding 20 days. The mechanisms underlying this inactivation are multifactorial, involving synergistic effects of elevated ammonia concentrations, volatile fatty acid accumulation, competition with indigenous microbiota, and elevated pH conditions
| [16] | Smith, S. R., Lang, N. L., Cheung, K. H. M., & Spanoudaki, K. (2005). Factors controlling pathogen destruction during anaerobic digestion of biowastes. Waste Management, 25(4), 417-425. https://doi.org/10.1016/j.wasman.2005.02.010 |
| [17] | Pandey, P. K., & Soupir, M. L. (2011). Escherichia coli inactivation kinetics in dairy manure under aerobic and anaerobic conditions. Bioresource Technology, 107, 204-211.
https://doi.org/10.1016/j.biortech.2011.12.017 |
[16, 17]
. In our study, the progressive pH increase from 6.5-6.8 to 8.8-9.4 likely contributed significantly to pathogen inactivation, as alkaline conditions disrupt cellular homeostasis in non-adapted bacteria
. Conversely, the persistence of
Bacillus spp. and
Clostridium spp. at detectable levels after 56 days, despite reductions of 3-5 log
10 CFU g
-1, underscores the inherent resilience of spore-forming bacteria to AD processes
. Spores are metabolically dormant structures encased in multiple protective layers that confer exceptional resistance to heat, desiccation, radiation, and chemical stress
. Bagge et al.
similarly observed that
Bacillus and
Clostridium species persisted throughout mesophilic AD, with complete inactivation requiring either thermophilic temperatures (>55°C) or extended retention times exceeding 90 days. In tropical contexts such as Côte d'Ivoire, where ambient temperatures fluctuate between 25°C and 37°C, achieving thermophilic conditions without external heating is economically challenging, rendering complete spore inactivation difficult to attain.
One of the most alarming findings of this study is the high prevalence of antimicrobial resistance among bacterial isolates recovered from pig slurry, with 75% of bacterial groups exhibiting MAR indices exceeding 0.2. This threshold, established by Krumperman
| [27] | Krumperman, P. H. (1983). Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Applied and Environmental Microbiology, 46(1), 165-170.
https://doi.org/10.1128/aem.46.1.165-170.1983 |
[27]
, distinguishes bacteria originating from environments with high antimicrobial selective pressure-such as intensive livestock operations-from those arising from pristine environments. The mean MAR index of 0.47 observed in our study substantially exceeds values typically reported from non-intensive farming systems and is comparable to indices documented in concentrated animal feeding operations (CAFOs) where antimicrobials are routinely administered for growth promotion and disease prophylaxis
| [10] | Manyi-Loh, C., Mamphweli, S., Meyer, E., & Okoh, A. (2018). Antibiotic use in agriculture and its consequential resistance in environmental sources: Potential public health implications. Molecules, 23(4), 795.
https://doi.org/10.3390/molecules23040795 |
[10]
.
The extraordinarily high resistance rates observed in spore-forming bacteria-
Clostridium spp. (66.7%) and
Bacillus spp. (64.3%)-warrant particular attention. These organisms not only survive AD but also carry multidrug resistance determinants that can persist in soil following digestate application
| [20] | Venglovsky, J., Sasakova, N., & Placha, I. (2009). Pathogens and antibiotic residues in animal manures and hygienic and ecological risks related to subsequent land application. Bioresource Technology, 100(22), 5386-5391.
https://doi.org/10.1016/j.biortech.2009.03.068 |
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[20, 22]
. Berendonk
et al. | [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[22]
demonstrated that agricultural soils amended with antibiotic-resistant bacteria serve as long-term reservoirs for resistance genes, which can subsequently transfer to indigenous soil microbiota and potentially to crop-associated or human-pathogenic bacteria via horizontal gene transfer mechanisms. Furthermore,
Clostridium difficile, which exhibited the highest MAR index (0.67) among spore-formers in our study, is an established nosocomial pathogen causing severe healthcare-associated infections globally
. Its persistence in digestate represents a direct public health hazard, particularly in regions with limited sanitation infrastructure where person-to-environment transmission pathways are more probable.
Intriguingly,
Enterobacteriaceae and
Salmonella spp. displayed remarkably low resistance rates (6.7% and 13.3%, respectively), with MAR indices well below the 0.2 threshold. This pattern suggests heterogeneity in antimicrobial exposure across different bacterial taxa within the same pig production systems. Several hypotheses may explain this observation. First, the pig farms supplying manure to the biodigesters may have selectively used antimicrobials targeting Gram-positive pathogens, exerting minimal selective pressure on Gram-negative bacteria. Second,
Enterobacteriaceae and
Salmonella populations in pig intestinal microbiota may experience higher turnover rates, limiting opportunities for resistance gene acquisition and fixation
| [10] | Manyi-Loh, C., Mamphweli, S., Meyer, E., & Okoh, A. (2018). Antibiotic use in agriculture and its consequential resistance in environmental sources: Potential public health implications. Molecules, 23(4), 795.
https://doi.org/10.3390/molecules23040795 |
[10]
. Third, these bacteria may harbor resistance genes that are phenotypically silent under standard laboratory testing conditions but could be mobilized under environmental stress
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[22]
.
A striking finding from our statistical analysis is the absence of correlation between MAR indices and bacterial persistence through AD (r = 0.23, p = 0.58). This result challenges the hypothesis that antibiotic resistance confers survival advantages during anaerobic digestion and instead underscores the primacy of physiological adaptations-particularly sporulation-in determining bacterial fate during waste treatment
. Sporulation is a complex developmental process triggered by nutrient limitation and environmental stress, enabling bacteria to enter a dormant state that is impervious to most antimicrobial agents and physical stressors
. Consequently, the ability to form spores, rather than possession of acquired resistance genes, emerges as the critical determinant of bacterial survival in AD systems.
Nevertheless, the co-occurrence of sporulation capacity and high-level antimicrobial resistance in
Bacillus and
Clostridium isolates presents a synergistic threat. These bacteria not only withstand the physical and chemical stresses of AD but also harbor extensive resistance repertoires that can disseminate into agricultural ecosystems via digestate land application
| [20] | Venglovsky, J., Sasakova, N., & Placha, I. (2009). Pathogens and antibiotic residues in animal manures and hygienic and ecological risks related to subsequent land application. Bioresource Technology, 100(22), 5386-5391.
https://doi.org/10.1016/j.biortech.2009.03.068 |
| [21] | Martens, W., & Böhm, R. (2009). Overview of the ability of different treatment methods for liquid and solid manure to inactivate pathogens. Bioresource Technology, 100(22), 5374-5378. https://doi.org/10.1016/j.biortech.2009.01.014 |
[20, 21]
. Venglovsky et al.
| [20] | Venglovsky, J., Sasakova, N., & Placha, I. (2009). Pathogens and antibiotic residues in animal manures and hygienic and ecological risks related to subsequent land application. Bioresource Technology, 100(22), 5386-5391.
https://doi.org/10.1016/j.biortech.2009.03.068 |
[20]
documented the long-term persistence of antibiotic resistance genes in soils receiving manure-based fertilizers, with detectable resistance determinants remaining present for months to years following application. Such environmental reservoirs facilitate resistance gene exchange among soil microbiota, creating opportunities for resistance transfer to plant pathogens, zoonotic agents, or environmental opportunists that can subsequently colonize humans or animals
| [8] | Walsh, F., & Duffy, B. (2013). The culturable soil antibiotic resistome: A community of multi-drug resistant bacteria. PLoS ONE, 8(6), e65567.
https://doi.org/10.1371/journal.pone.0065567 |
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[8, 22]
.
This study exemplifies the interconnectedness of animal health, environmental health, and human health-the core tenets of the One Health framework
. Intensive livestock production systems, driven by the need to meet growing global protein demands, frequently rely on antimicrobials for disease control and growth enhancement
| [1] | Muylaert, K., Bastiaens, L., Vandamme, D., & Gouveia, L. (2017). Harvesting of microalgae: Overview of process options and their strengths and drawbacks. Microalgae-based biofuels and bioproducts, 113-132.
https://doi.org/10.1016/B978-0-08-101023-5.00005-4 |
| [2] | Ribeiro, C. F. A., Silveira, G. G. D. O. S., Candido, E. D. S., Cardoso, M. H., Espinola Carvalho, C. M., & Franco, O. L. (2020). Effects of antibiotic treatment on gut microbiota and how to overcome its negative impacts on human health. ACS Infectious Diseases, 6(10), 2544-2559.
https://doi.org/10.1021/acsinfecdis.0c00036 |
| [10] | Manyi-Loh, C., Mamphweli, S., Meyer, E., & Okoh, A. (2018). Antibiotic use in agriculture and its consequential resistance in environmental sources: Potential public health implications. Molecules, 23(4), 795.
https://doi.org/10.3390/molecules23040795 |
[1, 2, 10]
. These practices generate large volumes of manure contaminated with antimicrobial residues, resistant bacteria, and mobile genetic elements encoding resistance
| [7] | Ouedraogo, A. S., Jean Pierre, H., Bañuls, A. L., Ouédraogo, R., & Godreuil, S. (2017). Emergence and spread of antibiotic resistance in West Africa: Contributing factors and threat assessment. Médecine et Santé Tropicales, 27(2), 147-154.
https://doi.org/10.1684/mst.2017.0678 |
| [8] | Walsh, F., & Duffy, B. (2013). The culturable soil antibiotic resistome: A community of multi-drug resistant bacteria. PLoS ONE, 8(6), e65567.
https://doi.org/10.1371/journal.pone.0065567 |
| [11] | Daghrir, R., & Drogui, P. (2013). Tetracycline antibiotics in the environment: A review. Environmental Chemistry Letters, 11(3), 209-227. https://doi.org/10.1007/s10311-013-0404-8 |
[7, 8, 11]
. When such manure is processed through AD and subsequently applied to agricultural land as digestate, it creates multiple exposure pathways through which resistant bacteria can reach humans: direct contact during farming activities, consumption of contaminated crops, groundwater infiltration, or aerosolization during field application
| [12] | Sharma, B., Dangi, A. K., & Shukla, P. (2016). Contemporary enzyme based technologies for bioremediation: A review. Journal of Environmental Management, 210, 10-22.
https://doi.org/10.1016/j.jenvman.2017.12.075 |
| [13] | Baquero, F., Martínez, J. L., & Cantón, R. (2008). Antibiotics and antibiotic resistance in water environments. Current Opinion in Biotechnology, 19(3), 260-265.
https://doi.org/10.1016/j.copbio.2008.05.006 |
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[12, 13, 22]
.
From a circular economy perspective, AD represents a sustainable approach to nutrient recycling and renewable energy generation, converting organic waste into biogas and nutrient-rich biofertilizer
| [14] | Okeke, E. S., Ita, R. E., Okoye, C. O., Ezeorba, T. P. C., & Mao, G. (2020). A critical review of life cycle assessment studies of renewable diesel fuels. Energies, 13(19), 5137.
https://doi.org/10.3390/en13195137 |
| [15] | Weiland, P. (2010). Biogas production: Current state and perspectives. Applied Microbiology and Biotechnology, 85(4), 849-860. https://doi.org/10.1007/s00253-009-2246-7 |
[14, 15]
. However, the findings of this study underscore that circular agricultural systems must incorporate antimicrobial resistance considerations into their design and operation to prevent inadvertent propagation of resistance through environmental pathways
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[22]
. As noted by Berendonk et al.
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[22]
, the environment functions as both a reservoir and transmission route for antimicrobial resistance, necessitating integrated surveillance and mitigation strategies that span agricultural, environmental, and clinical domains.
In tropical developing countries such as Côte d'Ivoire, where this study was conducted, the challenges are particularly acute. Limited regulatory oversight of antimicrobial use in livestock production, inadequate waste management infrastructure, and resource constraints impede the implementation of comprehensive AMR control measures
| [7] | Ouedraogo, A. S., Jean Pierre, H., Bañuls, A. L., Ouédraogo, R., & Godreuil, S. (2017). Emergence and spread of antibiotic resistance in West Africa: Contributing factors and threat assessment. Médecine et Santé Tropicales, 27(2), 147-154.
https://doi.org/10.1684/mst.2017.0678 |
[7]
. Nevertheless, the rapid expansion of pig production in West Africa-driven by increasing urbanization and protein consumption-makes these issues increasingly urgent. Our findings from Abidjan pig farms reflect broader patterns likely occurring across the region, highlighting the need for region-specific interventions tailored to local socioeconomic and environmental contexts.
Several limitations of this study merit acknowledgment. First, our investigation focused exclusively on culturable bacteria, potentially underestimating the true diversity and abundance of antimicrobial-resistant organisms present in pig slurry and digestate. Molecular approaches such as metagenomics and quantitative PCR targeting resistance genes would provide more comprehensive surveillance of the resistome
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[22]
. Second, we characterized phenotypic resistance profiles but did not identify the specific genetic mechanisms underlying observed resistance, precluding assessment of resistance gene transfer potential. Third, the study was conducted at pilot scale under tropical ambient conditions; findings may not directly extrapolate to full-scale thermophilic digesters or temperate climates where temperature dynamics differ substantially.
Future research should prioritize several key areas. Molecular characterization of resistance genes using whole-genome sequencing would elucidate the genetic basis of observed resistance and identify mobile genetic elements facilitating resistance dissemination
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[22]
. Longitudinal field studies tracking resistant bacteria and resistance genes in soils following digestate application would quantify environmental persistence and assess transmission risks to crops and groundwater. Additionally, techno-economic analyses evaluating post-AD treatment technologies-such as composting, pasteurization, or advanced oxidation-would inform practical strategies for enhancing digestate safety without compromising economic viability. Finally, integrated surveillance systems linking livestock production, waste treatment, environmental reservoirs, and clinical settings would enable comprehensive One Health monitoring of AMR emergence and dissemination pathways
| [3] | World Health Organization. (2015). Global action plan on antimicrobial resistance. WHO Press.
https://www.who.int/publications/i/item/9789241509763 |
| [6] | World Health Organization. (2018). World health statistics 2018: Monitoring health for the SDGs, sustainable development goals. WHO Press.
https://www.who.int/publications/i/item/9789241565585 |
| [22] | Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., Bürgmann, H., Sørum, H., Norström, M., Pons, M. N., Kreuzinger, N., Huovinen, P., Stefani, S., Schwartz, T., Kisand, V., Baquero, F., & Martinez, J. L. (2015). Tackling antibiotic resistance: The environmental framework. Nature Reviews Microbiology, 13(5), 310-317.
https://doi.org/10.1038/nrmicro3439 |
[3, 6, 22]
.
5. Conclusion
This study provides comprehensive evidence that mesophilic anaerobic digestion of pig slurry effectively reduces most pathogenic bacteria but incompletely eliminates spore-forming antibiotic-resistant organisms. While non-spore-formers such as E. coli, Enterococcus, and Salmonella were completely inactivated within 56 days, Bacillus and Clostridium species persisted despite 3-5 log10 reductions. Alarmingly, six of eight bacterial groups exhibited MAR indices exceeding 0.2, indicating high-risk antimicrobial contamination sources. The persistence of spore-forming multidrug-resistant bacteria in digestate poses significant public health risks when applied to agricultural land, particularly in tropical developing regions with limited regulatory oversight.
From a One Health perspective, these findings underscore the urgency of integrating antimicrobial resistance considerations into circular economy waste management systems. Sustainable solutions require multifaceted interventions spanning judicious antimicrobial stewardship in livestock production, optimization of AD technologies to enhance pathogen inactivation, implementation of complementary post-treatment measures (composting, pasteurization), and establishment of integrated surveillance systems monitoring resistance emergence across agricultural, environmental, and clinical domains. In tropical contexts where thermophilic AD is economically constrained, alternative hygienization strategies-such as prolonged retention times, alkaline stabilization, or sequential treatment processes-merit exploration.
As global livestock production intensifies to meet rising protein demands, the environmental dimensions of antimicrobial resistance will become increasingly critical. This study from Abidjan, Côte d'Ivoire, contributes to the growing body of evidence demonstrating that agricultural waste management practices function as critical nodes in AMR transmission networks. Addressing this challenge requires transdisciplinary collaboration among veterinarians, agronomists, environmental scientists, microbiologists, and public health professionals, working within a One Health framework to develop context-appropriate, economically viable, and environmentally sustainable solutions for mitigating antimicrobial resistance in circular agricultural systems.