Ecological restoration partially relies on plant regeneration from the above-ground vegetation (AGV) and soil seed banks (SSB); however, the co-occurrence of species in these sources vary across ecosystems. In this study, we measured soil properties and surveyed SSB and AGV, monitored regenerants for two years to estimate survival rates. We assessed the variation of species abundance in AGV, SSB and natural regenerants using a log-linear model and tested for correlation between soil properties using Pearson’s correlation coefficient and the association between the importance value index and survival rate of regenerants using generalised linear models. We found within-community dissimilarity between AGV and SSB, and the co-occurrence of species was predicted by periods of invasive species removal. There were five (4.46%) common species and five (4.46%), 34 (30.36%) and 42 (37.5%) exclusive species in natural regenerants, SSB and AGV, respectively. The common species among all periods of invasive species removal were 12.5%, 20%, and 41.7% in AGV, natural regenerants, and SSB, respectively. The diversity of SSB was positively associated with the survival rate of regenerants, but not AGV diversity. Survival rates of regenerants were associated with seed mass but not plant height, while plant functional traits were not associated with importance value indices of AGV. These findings show that native plant recovery can be accelerated by removal of invasive species; however, diversity differences exist between AGV and SSB, which were not necessarily modulated by plant functional traits and soil characteristics.
| Published in | Journal of Plant Sciences (Volume 14, Issue 2) |
| DOI | 10.11648/j.jps.20261402.12 |
| Page(s) | 79-92 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Afromontane Forest, Ecosystem Recovery, Ecosystem Restoration, Invasive Species, Natural Regenerants, Soil Seed Bank
Traits | Predictor | Importance value index (IVI) | Plant survival rate | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
Estimate | SE | statistic | pvalue | AIC | Estimate | SE | statistic | pvalue | AIC | ||
Growth form | (Intercept) | 7.758 | 5.14 | 1.509 | 0.139 | 287.32 | -0.299 | 0.56 | -0.529 | 0.603 | 61.08 |
Plant height | 0.084 | 0.10 | 0.834 | 0.409 | 0.002 | 0.02 | 0.067 | 0.948 | |||
Seed mass | -0.443 | 0.32 | -1.377 | 0.176 | 1.264 | 0.44 | 2.852 | 0.011* | |||
Shrub | -5.594 | 5.30 | -1.055 | 0.298 | -0.282 | 0.69 | -0.408 | 0.688 | |||
Tree | -3.116 | 5.38 | -0.580 | 0.565 | 0.345 | 0.74 | 0.466 | 0.647 | |||
Fruit type | (Intercept) | -3.656 | 4.55 | -0.804 | 0.429 | 195.83 | -0.386 | 0.95 | -0.406 | 0.694 | 38.40 |
Plant height | 0.264 | 0.13 | 1.977 | 0.059 | 0.016 | 0.03 | 0.546 | 0.598 | |||
Seed mass | -0.403 | 0.39 | -1.046 | 0.305 | 1.503 | 0.68 | 2.224 | 0.053 | |||
Biotic | 6.395 | 3.45 | 1.860 | 0.074 | -0.031 | 0.92 | -0.034 | 0.973 | |||
Deciduousness | (Intercept) | 6.054 | 2.43 | 2.492 | 0.018* | 223.57 | -0.496 | 0.42 | -1.183 | 0.251 | 60.25 |
Plant height | 0.133 | 0.09 | 1.420 | 0.166 | 0.017 | 0.02 | 0.989 | 0.335 | |||
Seed mass | -0.321 | 0.35 | -0.920 | 0.365 | 1.354 | 0.43 | 3.152 | 0.005* | |||
Evergreen | -3.379 | 2.09 | -1.621 | 0.115 | 0.186 | 0.36 | 0.512 | 0.615 | |||
Seed dispersal | (Intercept) | 1.582 | 3.67 | 0.431 | 0.671 | 181.16 | -0.949 | 0.77 | -1.230 | 0.235 | 60.12 |
Plant height | 0.163 | 0.09 | 1.703 | 0.102 | 0.005 | 0.02 | 0.285 | 0.779 | |||
Seed mass | -0.295 | 0.33 | -0.907 | 0.374 | 1.228 | 0.42 | 2.923 | 0.009* | |||
Autochorous | 13.256 | 4.86 | 2.730 | 0.012* | 0.179 | 0.89 | 0.199 | 0.845 | |||
Unspecialized | -0.457 | 6.04 | -0.076 | 0.940 | 0.379 | 1.09 | 0.349 | 0.732 | |||
Zoochorous | 1.409 | 3.58 | 0.393 | 0.698 | 0.981 | 0.83 | 1.183 | 0.253 | |||
AGV | Above-Ground Vegetation |
AIC | Akaike Information Criterion |
BBGF | Brackenhurst Botanical Garden and Forest |
CCA | Canonical Correspondence Analysis |
DBH | Diameter at Breast Height |
GIFT | Global Inventory of Floras and Traits |
GLM | Generalised Linear Model |
ISTA | International Seed Testing Association |
IVI | Importance Value Index |
KALRO | Kenya Agricultural & Livestock Research Organization |
LRT | Likelihood Ratio Test |
RCD | Root Collar Diameter |
RD | Relative Density |
RDO | Relative Dominance |
RF | Relative Frequency |
SE | Standard Error |
SER | Society for Ecological Restoration |
SSB | Soil Seed Bank |
| [1] | Al-Huqail, A. A., Al-Harbi, H. F., Alowaifeer, A. M., El-Sheikh, M. A., Assaeed, A. M., Alsaleem, T. S., Kassem, H. S., Azab, O. M., Dar, B. A., Malik, J. A., & Abd-ElGawad, A. M. (2025). Correlation between aboveground vegetation composition and soil seed bank of Raudhat desert habitat: A case study of Raudhat Alkhafs, Saudi Arabia. BMC Plant Biology, 25(1), 136. |
| [2] | Anju, V. M., Warrier, R. R., & Kunhikannan, C. (2022). Significance of soil seed bank in forest vegetation—A review. Seeds, 1(3), 181–197. |
| [3] | Ashiono, F., Kamiri, H., & Kinyanjui, M. (n.d.). Evaluation of mineral nutrition and growth of Eucalyptus saligna seedlings raised on organic-enriched nursery potting media. Journal of Research in Forestry, Wildlife & Environment, 11, 39–50. |
| [4] | Bekele, M., Demissew, S., Bekele, T., & Woldeyes, F. (2022). Soil seed bank distribution and restoration potential in the vegetation of Buska Mountain range, Hamar district, southwestern Ethiopia. Heliyon, 8(11), e11244. |
| [5] | Birhanu, L., Bekele, T., Tesfaw, B., & Demissew, S. (2022). Soil seed bank composition and aboveground vegetation in dry Afromontane forest patches of Northwestern Ethiopia. Trees, Forests and People, 9, 100292. |
| [6] | Brandt, A. J., Png, G. K., Jo, I., McGrannachan, C., Allen, K., Peltzer, D. A., D’Antonio, C., Dickie, I. A., French, K., Leishman, M. R., Ostertag, R., Parker, I. M., Stanley, M. C., Suding, K. N., & Bellingham, P. J. (2023). Managing multi-species plant invasions when interactions influence their impact. Frontiers in Ecology and the Environment, 21(8), 370–379. |
| [7] | Buisson, E., Fidelis, A., Overbeck, G. E., Schmidt, I. B., Durigan, G., Young, T. P., Alvarado, S. T., Arruda, A. J., Boisson, S., Bond, W., Coutinho, A., Kirkman, K., Oliveira, R. S., Schmitt, M. H., Siebert, F., Siebert, S. J., Thompson, D. I., & Silveira, F. A. O. (2021). A research agenda for the restoration of tropical and subtropical grasslands and savannas. Restoration Ecology, 29(S1), e13292. |
| [8] | Chapman, C. A., Kitajima, K., Zanne, A. E., Kaufman, L. S., & Lawes, M. J. (2008). A 10-year evaluation of the functional basis for regeneration habitat preference of trees in an African evergreen forest. Forest Ecology and Management, 255(11), 3790–3796. |
| [9] | Chen, S., Hu, X., Baskin, C. C., & Baskin, J. M. (2024). A long-term experiment reveals no trade-off between seed persistence and seedling emergence. New Phytologist, 241(2), 623–631. |
| [10] | Dann, L. E., Guja, L., Kark, S., & Dwyer, J. (2024). Comparative study reveals management of a dominant invasive plant facilitates subtropical forest regeneration. Biological Invasions, 26(1), 299–313. |
| [11] | Denelle, P., Weigelt, P., & Kreft, H. (2023). gift—An r package to access the global inventory of floras and traits. Methods in Ecology and Evolution, 14(11), 2738–2748. |
| [12] | Dhakal, B., Pinard, M. A., Gunatilleke, I. A. U. N., Gunatilleke, C. V. S., & Burslem, D. F. R. P. (2015). Strategies for restoring tree seedling recruitment in high conservation value tropical montane forests underplanted with cardamom. Applied Vegetation Science, 18(1), 121–133. |
| [13] | Dyderski, M. K., & Jagodziński, A. M. (2020). Impact of invasive tree species on natural regeneration species composition, diversity, and density. Forests, 11(4), 456. |
| [14] | Egeta, D., Yadesa, G., Eshete, A., Belay, H., Charu, U., & Teshome, T. (2024). Woody species composition, structure, and soil seed bank of the degraded parts of the gera moist afromontane forest, southwestern ethiopia. International Journal of Forestry Research, 2024(1), 9849957. |
| [15] | Fabšičová, M., Vymyslický, T., Frei, I., Zdražílková, M., Smetanová, S., Winkler, J., & Jiroušek, M. (2024). The importance of soil seed banks for biodiversity restoration in degraded grasslands. Folia Geobotanica, 59(1), 17–37. |
| [16] | Flory, S. L., & Clay, K. (2009). Invasive plant removal method determines native plant community responses. Journal of Applied Ecology, 46(2), 434–442. |
| [17] | Gann, G. D., McDonald, T., Walder, B., Aronson, J., Nelson, C. R., Jonson, J., Hallett, J. G., Eisenberg, C., Guariguata, M. R., Liu, J., Hua, F., Echeverría, C., Gonzales, E., Shaw, N., Decleer, K., & Dixon, K. W. (2019). International principles and standards for the practice of ecological restoration. Second edition. Restoration Ecology, 27(S1). |
| [18] | Gioria, M., Carta, A., Baskin, C. C., Dawson, W., Essl, F., Kreft, H., Pergl, J., Van Kleunen, M., Weigelt, P., Winter, M., & Pyšek, P. (2021). Persistent soil seed banks promote naturalisation and invasiveness in flowering plants. Ecology Letters, 24(8), 1655–1667. |
| [19] | Guo, Y., Li, Y., Li, J., Li, J., Wen, S., Huang, F., He, W., Wang, B., Lu, S., Li, D., Xiang, W., & Li, X. (2022). Comparison of aboveground vegetation and soil seed bank composition among three typical vegetation types in the karst regions of southwest china. Agronomy, 12(8), 1871. |
| [20] | Hinkson, K. M., NeSmith, J. E., Alba, C., Durham, M., Ferrell, J., & Flory, S. L. (2024). Selective method for invasive plant removal enhances restoration. Restoration Ecology, 32(4), e14112. |
| [21] | ISTA. (2024). International Rules for Seed Testing: Sampling. International Rules for Seed Testing. International Rules for Seed Testing, 2024(1), 1–52. |
| [22] | Jakovac, C. C., Junqueira, A. B., Crouzeilles, R., Peña-Claros, M., Mesquita, R. C. G., & Bongers, F. (2021). The role of land-use history in driving successional pathways and its implications for the restoration of tropical forests. Biological Reviews, 96(4), 1114–1134. |
| [23] | Jennifer, N. M., J, I. K., R, M. C., & W, M. (2011). Assessment of soil seed bank from six different vegetation types in Kakamega forest, Western Kenya. African Journal of Biotechnology, 10(65), 14384–14391. |
| [24] | Kaigongi, M. M., Muturi, G. M., Kigomo, J. N., Gathara, M., & Otuoma, J. (2023). Mixed species natural forest regeneration trajectory in clear-felled monoculture plantation sites in Kenya: A step towards developing a natural forests restoration framework. African Journal of Ecology, 61(2), 513–518. |
| [25] | Karani, A., & Njoroge, L. (2025). Southernmost record of the threatened platycypha amboniensis martin 1915 (Odonata: Chlorocyphidae), endemic to the highlands of central kenya. Entomological Communications, 7, ec07015. |
| [26] | Kikoti, I. A., Ndangalasi, H. J., Kayombo, C. J., & Musoma, R. M. (2025). Soil seed bank composition and diversity in areas invaded with invasive alien species in the lower montane forest of kilimanjaro national park, tanzania. East African Journal of Environment and Natural Resources, 8(1), 195–218. |
| [27] | Kipkoech, S., Melly, D. K., Watuma Muema, B., Wei, N., Kamau, P., Kirika, P. M., Wang, Q., & Hu, G. (2020). An annotated checklist of the vascular plants of aberdare ranges forest, a part of eastern afromontane biodiversity hotspot. PhytoKeys, 149, 1–88. |
| [28] | Kovács-Hostyánszki, A., Szigeti, V., Miholcsa, Z., Sándor, D., Soltész, Z., Török, E., & Fenesi, A. (2022). Threats and benefits of invasive alien plant species on pollinators. Basic and Applied Ecology, 64, 89–102. |
| [29] | Lenda, M., Steudel, B., Skórka, P., Zagrodzka, Z. B., Moroń, D., Bączek-Kwinta, R., Janowiak, F., Baran, A., Possingham, H. P., & Knops, J. M. H. (2023). Multiple invasive species affect germination, growth, and photosynthesis of native weeds and crops in experiments. Scientific Reports, 13(1), 22146. |
| [30] | Macera, L. G., Pereira, S. R., & Souza, A. L. T. D. (2017). Survival and growth of tree seedlings as a function of seed size in a gallery forest under restoration. Acta Botanica Brasilica, 31(4), 539–545. |
| [31] | Makokha, J. (2018). Invasion of < i> Cestrum aurantiacum</i> Lindl. In Kenya. Journal of Environmental Protection, 09(06), 671–690. |
| [32] | Metz, J., Liancourt, P., Kigel, J., Harel, D., Sternberg, M., & Tielbörger, K. (2010). Plant survival in relation to seed size along environmental gradients: A long-term study from semi-arid and Mediterranean annual plant communities. Journal of Ecology, 98(3), 697–704. |
| [33] | Mganga, K. Z., Kaindi, E., Ndathi, A. J. N., Bosma, L., Kioko, T., Kadenyi, N., Wambua, S. M., Van Steenbergen, F., & Musimba, N. K. R. (2021). Morphoecological characteristics of grasses used to restore degraded semi-arid African rangelands. Ecological Solutions and Evidence, 2(2), e12078. |
| [34] | Miao, R., Song, Y., Sun, Z., Guo, M., Zhou, Z., & Liu, Y. (2016). Soil seed bank and plant community development in passive restoration of degraded sandy grasslands. Sustainability, 8(6), 581. |
| [35] | Mndela, M., Madakadze, C. I., Nherera-Chokuda, F., & Dube, S. (2020). Is the soil seed bank a reliable source for passive restoration of bush-cleared semi-arid rangelands of South Africa? Ecological Processes, 9(1), 1. |
| [36] | Mohammed, S. A., & Denboba, M. A. (2020). Study of soil seed banks in ex-closures for restoration of degraded lands in the central rift valley of ethiopia. Scientific Reports, 10(1), 956. |
| [37] | Mungi, N. A., Qureshi, Q., & Jhala, Y. V. (2023). Distribution, drivers and restoration priorities of plant invasions in India. Journal of Applied Ecology, 60(11), 2400–2412. |
| [38] | Nicholson, M. (2010). Brackenhurst Forest Botanic Garden, Kenya: Towards a self-sustaining botanic garden. Proceedings of the 4th Global Botanic Gardens Congress. |
| [39] | Ojija, F., Petruzzellis, F., & Bacaro, G. (2024). Review of invasive plant functional traits and management using remote sensing in sub-saharan africa. International Journal of Plant Biology, 15(2), 358–374. |
| [40] | Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R. B., Simpson, G. L., Solymos, P., Stevens, M. H. H., & Wagner, H. (2019). vegan: Community Ecology Package [R package version 2. 5-6. |
| [41] | Pedrini, S., & Dixon, K. W. (2020). International principles and standards for native seeds in ecological restoration. Restoration Ecology, 28(S3). |
| [42] | Perring, M. P., Standish, R. J., Price, J. N., Craig, M. D., Erickson, T. E., Ruthrof, K. X., Whiteley, A. S., Valentine, L. E., & Hobbs, R. J. (2015). Advances in restoration ecology: Rising to the challenges of the coming decades. Ecosphere, 6(8), 1–25. |
| [43] | Plohák, P., Švehláková, H., Stalmachová, B., Goňo, M., & Dvorský, T. (2025). Combining extraction and cultivation methods for soil seed bank analysis increases number of captured species and their similarity to above-ground vegetation. Frontiers in Plant Science, 15, 1500941. |
| [44] | Pyšek, P., Jarošík, V., Hulme, P. E., Pergl, J., Hejda, M., Schaffner, U., & Vilà, M. (2012). A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Global Change Biology, 18(5), 1725–1737. |
| [45] | R Core Team. (2024). _R: A Language and Environment for Statistical Computing_ [Computer software]. R Foundation for Statistical Computing. |
| [46] | Rahayu, S., Pambudi, S., Permadi, D., Tata, H. L., Martini, E., Rasnovi, S., Nuroniah, H. S., Kindt, R., Nugraha, M., Dewi, S., & Van Noordwijk, M. (2022). Functional trait profiles and diversity of trees regenerating in disturbed tropical forests and agroforests in Indonesia. Forest Ecosystems, 9, 100030. |
| [47] | Ricotta, C., & Szeidl, L. (2009). Diversity partitioning of Rao’s quadratic entropy. Theoretical Population Biology, 76(4), 299–302. |
| [48] | Rose, S., & Poorter, L. (2003). The importance of seed mass for early regeneration in tropical forest: A review. In Long-term changes in tropical tree diversity: Studies from the Guiana Shield, Africa, Borneo and Melanesia (pp. 19–35). Tropenbos International. |
| [49] | Ruwanza, S. (2021). Effects of solanum mauritianum scopoli (Bugweed) invasion on soil and vegetation in vhembe biosphere reserve, south africa. Austral Ecology, 46(3), 342–348. |
| [50] | Sanou, L., Zida, D., Savadogo, P., & Thiombiano, A. (2018). Comparison of aboveground vegetation and soil seed bank composition at sites of different grazing intensity around a savanna-woodland watering point in West Africa. Journal of Plant Research, 131(5), 773–788. |
| [51] | Shaw, K., Nicholson, M., & Hardwick, K. (2015). Encouraging and Enabling a Science-Based Approach to Ecological Restoration: An Introduction to the Work of the Ecological Restoration Alliance of Botanic Gardens (ERA). Sibbaldia: The International Journal of Botanic Garden Horticulture, 13, 145–152. |
| [52] | Shaw, K., Ruyonga, G., & Nicholson, M. (2016). Enhancing Tree Conservation and Forest Restoration in East Africa. BGjournal, 13, 28–31. |
| [53] | Spînu, A. P., Mysiak, W., Bauhus, J., Bielak, K., & Niklasson, M. (2023). Pioneer tree species accelerate restoration of tree-related microhabitats in 50-year-old reserves of Białowieża Forest, Poland. Ecology and Evolution, 13(7), e10238. |
| [54] | Tamura, A. (2016). Potential of soil seed banks in the ecological restoration of overgrazed floor vegetation in a cool-temperate old-growth damp forest in eastern Japan. Journal of Forest Research, 21(1), 43–56. |
| [55] | Tessema, Z. K., Ejigu, B., & Nigatu, L. (2017). Tree species determine soil seed bank composition and its similarity with understory vegetation in a semi-arid African savanna. Ecological Processes, 6(1), 9. |
| [56] | Toma, T. S. P., Overbeck, G. E., Mendonça, M. D. S., & Fernandes, G. W. (2023). Optimal references for ecological restoration: The need to protect references in the tropics. Perspectives in Ecology and Conservation, 21(1), 25–32. |
| [57] | Tóth, Á., Deák, B., Kelemen, A., Kiss, R., Lukács, K., Bátori, Z., & Valkó, O. (2025). Vertical stratification of the soil seed bank in wet grasslands and its implications for restoration. Community Ecology, 26(1), 131–142. |
| [58] | Ulyshen, M. D., Horn, S., & Hanula, J. L. (2022). Decadal patterns of forest and pollinator recovery following the eradication of an invasive shrub. Frontiers in Ecology and Evolution, 10, 832268. |
| [59] | Viani, R. A. G., Vidas, N. B., Pardi, M. M., Castro, D. C. V., Gusson, E., & Brancalion, P. H. S. (2015). Animal-dispersed pioneer trees enhance the early regeneration in Atlantic Forest restoration plantations. Natureza & Conservação, 13(1), 41–46. |
| [60] | Vujanović, D., Losapio, G., Milić, S., & Milić, D. (2022). The impact of multiple species invasion on soil and plant communities’ increases with invasive species co-occurrence. Frontiers in Plant Science, 13, 875824. |
| [61] | Walentowitz, A., Manthey, M., Bentet Preciado, M. B., Chango, R., Sevilla, C., & Jäger, H. (2021). Limited natural regeneration of unique Scalesia forest following invasive plant removal in Galapagos. PLOS ONE, 16(10), e0258467. |
| [62] | Weidlich, E. W. A., Flórido, F. G., Sorrini, T. B., & Brancalion, P. H. S. (2020). Controlling invasive plant species in ecological restoration: A global review. Journal of Applied Ecology, 57(9), 1806–1817. |
| [63] | Weigelt, P., König, C., & Kreft, H. (2020). Gift – a global inventory of floras and traits for macroecology and biogeography. Journal of Biogeography, 47(1), 16–43. |
| [64] | Witt, A., Beale, T., & Van Wilgen, B. W. (2018). An assessment of the distribution and potential ecological impacts of invasive alien plant species in eastern Africa. Transactions of the Royal Society of South Africa, 73(3), 217–236. |
| [65] | Wortley, L., Hero, J., & Howes, M. (2013). Evaluating ecological restoration success: A review of the literature. Restoration Ecology, 21(5), 537–543. |
| [66] | Zenni, R. D., Da Cunha, W. L., Musso, C., De Souza, J. V., Nardoto, G. B., & Miranda, H. S. (2020). Synergistic impacts of co-occurring invasive grasses cause persistent effects in the soil-plant system after selective removal. Functional Ecology, 34(5), 1102–1112. |
| [67] | Zhou, Y., Liu, B., Mbuni, Y., Yan, X., Mwachala, G., Hu, G., & Wang, Q. (2017). Vascular flora of kenya, based on the flora of tropical east africa. PhytoKeys, 90, 113–126. |
APA Style
Karani, A., Gichira, A., Musembi, T., Adhiambo, L., Otieno, V., et al. (2026). Implications of Invasive Species Removal, Soil Properties and Plant Functional Traits on Survival and Co-occurrence of Above-ground Vegetation with Soil Seed Bank. Journal of Plant Sciences, 14(2), 79-92. https://doi.org/10.11648/j.jps.20261402.12
ACS Style
Karani, A.; Gichira, A.; Musembi, T.; Adhiambo, L.; Otieno, V., et al. Implications of Invasive Species Removal, Soil Properties and Plant Functional Traits on Survival and Co-occurrence of Above-ground Vegetation with Soil Seed Bank. J. Plant Sci. 2026, 14(2), 79-92. doi: 10.11648/j.jps.20261402.12
AMA Style
Karani A, Gichira A, Musembi T, Adhiambo L, Otieno V, et al. Implications of Invasive Species Removal, Soil Properties and Plant Functional Traits on Survival and Co-occurrence of Above-ground Vegetation with Soil Seed Bank. J Plant Sci. 2026;14(2):79-92. doi: 10.11648/j.jps.20261402.12
@article{10.11648/j.jps.20261402.12,
author = {Anthony Karani and Andrew Gichira and Tobin Musembi and Lucy Adhiambo and Victor Otieno and Jonathan Jenkins},
title = {Implications of Invasive Species Removal, Soil Properties and Plant Functional Traits on Survival and Co-occurrence of Above-ground Vegetation with Soil Seed Bank},
journal = {Journal of Plant Sciences},
volume = {14},
number = {2},
pages = {79-92},
doi = {10.11648/j.jps.20261402.12},
url = {https://doi.org/10.11648/j.jps.20261402.12},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.jps.20261402.12},
abstract = {Ecological restoration partially relies on plant regeneration from the above-ground vegetation (AGV) and soil seed banks (SSB); however, the co-occurrence of species in these sources vary across ecosystems. In this study, we measured soil properties and surveyed SSB and AGV, monitored regenerants for two years to estimate survival rates. We assessed the variation of species abundance in AGV, SSB and natural regenerants using a log-linear model and tested for correlation between soil properties using Pearson’s correlation coefficient and the association between the importance value index and survival rate of regenerants using generalised linear models. We found within-community dissimilarity between AGV and SSB, and the co-occurrence of species was predicted by periods of invasive species removal. There were five (4.46%) common species and five (4.46%), 34 (30.36%) and 42 (37.5%) exclusive species in natural regenerants, SSB and AGV, respectively. The common species among all periods of invasive species removal were 12.5%, 20%, and 41.7% in AGV, natural regenerants, and SSB, respectively. The diversity of SSB was positively associated with the survival rate of regenerants, but not AGV diversity. Survival rates of regenerants were associated with seed mass but not plant height, while plant functional traits were not associated with importance value indices of AGV. These findings show that native plant recovery can be accelerated by removal of invasive species; however, diversity differences exist between AGV and SSB, which were not necessarily modulated by plant functional traits and soil characteristics.},
year = {2026}
}
TY - JOUR T1 - Implications of Invasive Species Removal, Soil Properties and Plant Functional Traits on Survival and Co-occurrence of Above-ground Vegetation with Soil Seed Bank AU - Anthony Karani AU - Andrew Gichira AU - Tobin Musembi AU - Lucy Adhiambo AU - Victor Otieno AU - Jonathan Jenkins Y1 - 2026/03/26 PY - 2026 N1 - https://doi.org/10.11648/j.jps.20261402.12 DO - 10.11648/j.jps.20261402.12 T2 - Journal of Plant Sciences JF - Journal of Plant Sciences JO - Journal of Plant Sciences SP - 79 EP - 92 PB - Science Publishing Group SN - 2331-0731 UR - https://doi.org/10.11648/j.jps.20261402.12 AB - Ecological restoration partially relies on plant regeneration from the above-ground vegetation (AGV) and soil seed banks (SSB); however, the co-occurrence of species in these sources vary across ecosystems. In this study, we measured soil properties and surveyed SSB and AGV, monitored regenerants for two years to estimate survival rates. We assessed the variation of species abundance in AGV, SSB and natural regenerants using a log-linear model and tested for correlation between soil properties using Pearson’s correlation coefficient and the association between the importance value index and survival rate of regenerants using generalised linear models. We found within-community dissimilarity between AGV and SSB, and the co-occurrence of species was predicted by periods of invasive species removal. There were five (4.46%) common species and five (4.46%), 34 (30.36%) and 42 (37.5%) exclusive species in natural regenerants, SSB and AGV, respectively. The common species among all periods of invasive species removal were 12.5%, 20%, and 41.7% in AGV, natural regenerants, and SSB, respectively. The diversity of SSB was positively associated with the survival rate of regenerants, but not AGV diversity. Survival rates of regenerants were associated with seed mass but not plant height, while plant functional traits were not associated with importance value indices of AGV. These findings show that native plant recovery can be accelerated by removal of invasive species; however, diversity differences exist between AGV and SSB, which were not necessarily modulated by plant functional traits and soil characteristics. VL - 14 IS - 2 ER -
Centre for Ecosystem Restoration, Limuru, Kenya;Department of Biological Sciences, University of Embu, Embu, Kenya
Centre for Ecosystem Restoration, Limuru, Kenya
Centre for Ecosystem Restoration, Limuru, Kenya
Centre for Ecosystem Restoration, Limuru, Kenya
Centre for Ecosystem Restoration, Limuru, Kenya
Centre for Ecosystem Restoration, Limuru, Kenya
Figure 1. Map of Kiambu County, showing sampling sites at the Brackenhurst Botanical Garden and Forest (BBGF) in Tigoni, Central Highlands of Kenya. Inset is the map of Kenya.
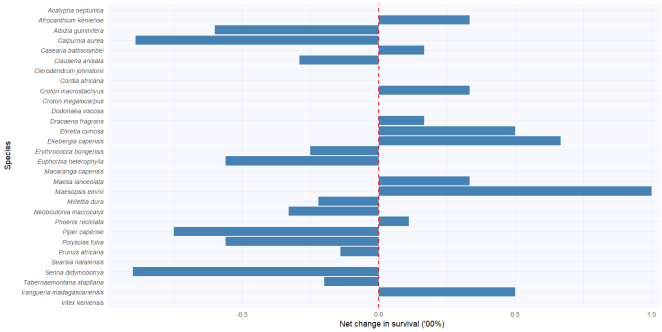
Figure 2. Bar graph showing plant species and net survival rate, expressed as the change in the number of regenerants across the study period. The zero intercept line represents no net change in abundance (100% survival rate); the positive scale indicates a net increase in the number of regenerants, while the negative scale indicates a net decrease.
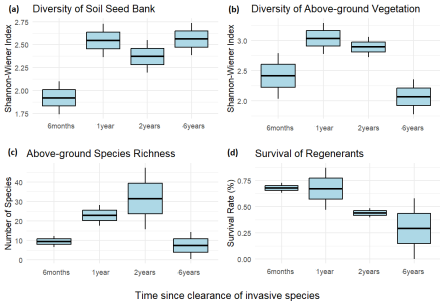
Figure 3. Box plot of different vegetation metrics, showing a) Shannon diversity of soil seed bank, b) Shannon diversity of above-ground vegetation, c) species richness of above-ground vegetation and average survival rate of regenerants. The box represents the interquartile range, the horizontal line is the median, and the whiskers extend to the smallest and largest values.
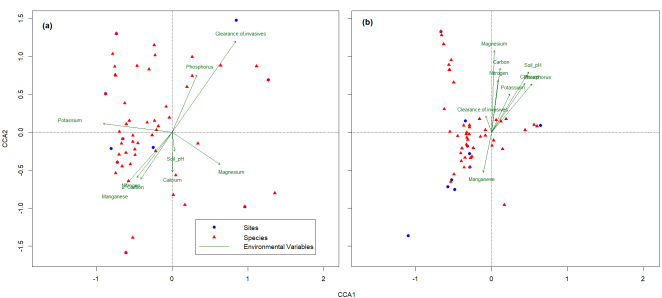
Figure 4. Canonical Correspondence Analysis (CCA) ordination biplot showing the relationship of plant composition (red triangles), sampling sites (blue dots), and soil properties for, (a) above-ground vegetation, and (b) soil seed bank. The arrows point at their respective centroids; the arrow direction indicates the relationship.
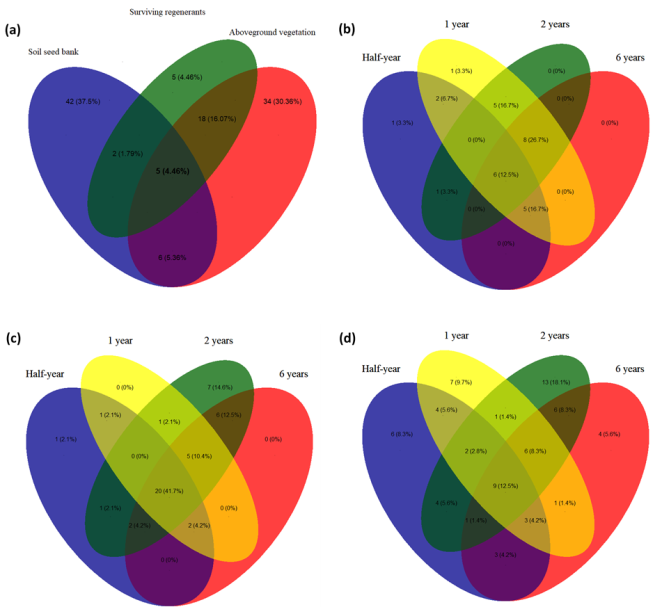
Figure 5. Venn diagram showing common and exclusive species in a) soil seed bank, above-ground vegetation, and regenerants and time since removal of invasive species in b) regenerants, c) soil seed bank, and d) above-ground vegetation.
Information

